

An allosteric propofol-binding site in kinesin disrupts kinesin-mediated processive movement on microtubules
- PMID: 29844014
- PMCID: PMC6065180
- DOI: 10.1074/jbc.RA118.002182
An allosteric propofol-binding site in kinesin disrupts kinesin-mediated processive movement on microtubules
Abstract
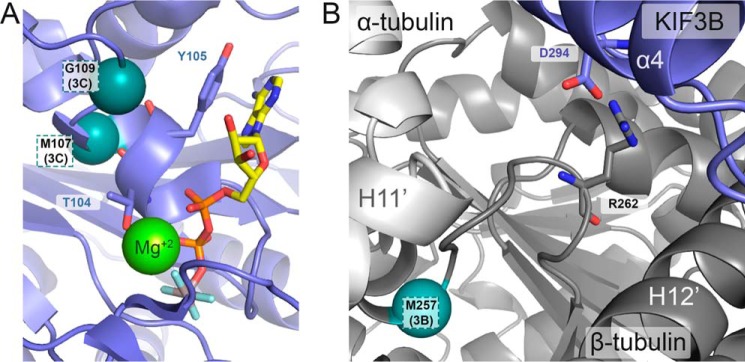
Microtubule-based molecular motors mediate transport of intracellular cargo to subdomains in neurons. Previous evidence has suggested that the anesthetic propofol decreases the average run-length potential of the major anterograde transporters kinesin-1 and kinesin-2 without altering their velocity. This effect on kinesin has not been observed with other inhibitors, stimulating considerable interest in the underlying mechanism. Here, we used a photoactive derivative of propofol, meta-azipropofol (AziPm), to search for potential propofol-binding sites in kinesin. Single-molecule motility assays confirmed that AziPm and propofol similarly inhibit kinesin-1 and kinesin-2. We then applied AziPm in semiquantitative radiolabeling and MS microsequencing assays to identify propofol-binding sites within microtubule-kinesin complexes. The radiolabeling experiments suggested preferential AziPm binding to the ATP-bound microtubule-kinesin complex. The photolabeled residues were contained within the kinesin motor domain rather than at the motor domain-β-tubulin interface. No residues within the P-loop of kinesin were photolabeled, indicating an inhibitory mechanism that does not directly affect ATPase activity and has an effect on run length without changing velocity. Our results also indicated that when the kinesin motor interacts with the microtubule during its processive run, a site forms in kinesin to which propofol can then bind and allosterically disrupt the kinesin-microtubule interaction, resulting in kinesin detachment and run termination. The discovery of the propofol-binding allosteric site in kinesin may improve our understanding of the strict coordination of the motor heads during the processive run. We hypothesize that propofol's potent effect on intracellular transport contributes to various components of its anesthetic action.
Keywords: anesthesia; kinesin; microtubule; molecular motor; photoaffinity labeling; propofol; single molecule motility; single-molecule biophysics; tubulin.
© 2018 Woll et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
