

Riboswitches and Translation Control
- PMID: 29844057
- PMCID: PMC6211393
- DOI: 10.1101/cshperspect.a032797
Riboswitches and Translation Control
Abstract
A growing collection of bacterial riboswitch classes is being discovered that sense central metabolites, coenzymes, and signaling molecules. Included among the various mechanisms of gene regulation exploited by these RNA regulatory elements are several that modulate messenger RNA (mRNA) translation. In this review, the mechanisms of riboswitch-mediated translation control are summarized to highlight both their diversity and potential ancient origins. These mechanisms include ligand-gated presentation or occlusion of ribosome-binding sites, control of alternative splicing of mRNAs, and the regulation of mRNA stability. Moreover, speculation on the potential for novel riboswitch discoveries is presented, including a discussion on the potential for the discovery of a greater diversity of mechanisms for translation control.
Copyright © 2018 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
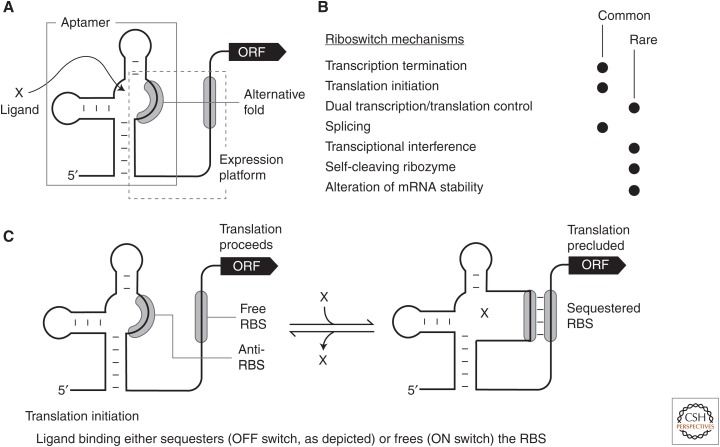
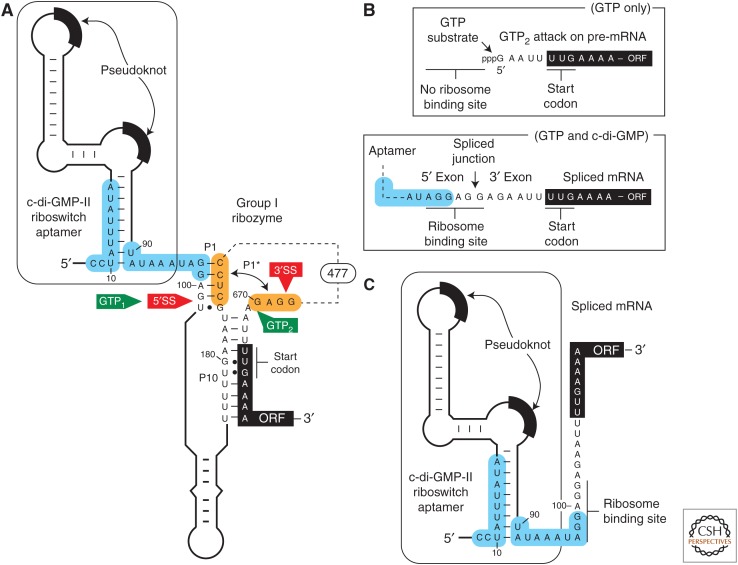
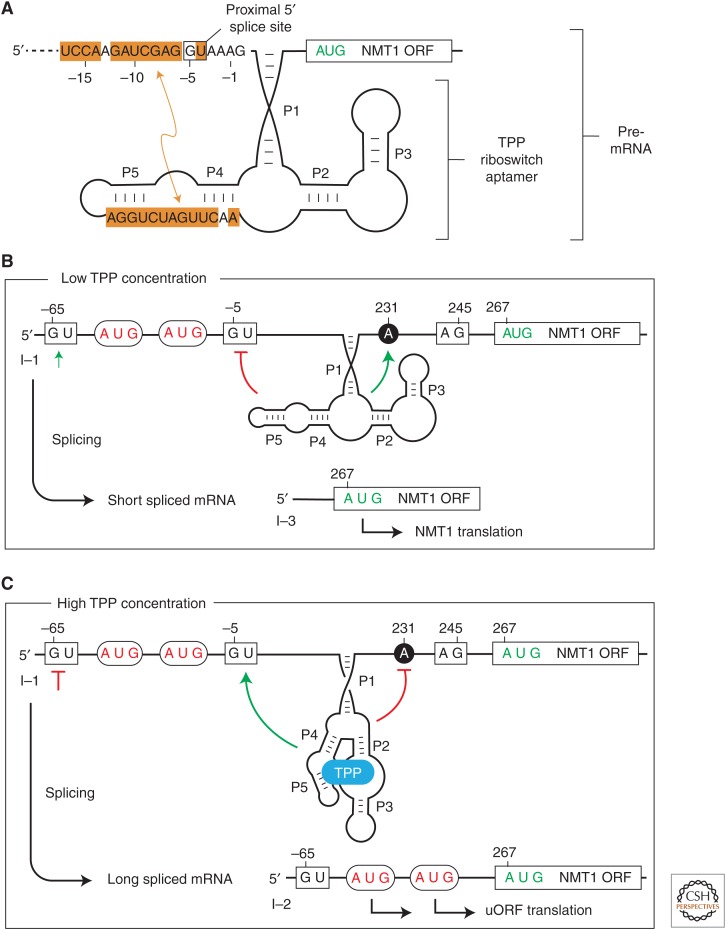
References
-
- Ames TD, Breaker RR. 2010. Bacterial riboswitch diversity and analysis. In The chemical biology of nucleic acids (ed. Mayer G), Chap. 20 Wiley, Chichester, UK.
-
- Antson AA, Dodson EJ, Dodson G, Greaves RB, Chen X, Gollnick P. 1999. Structure of the trp RNA-binding attenuation protein, TRAP, bound to RNA. Nature 401: 235–242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources