

Fungal Isocyanide Synthases and Xanthocillin Biosynthesis in Aspergillus fumigatus
- PMID: 29844112
- PMCID: PMC5974471
- DOI: 10.1128/mBio.00785-18
Fungal Isocyanide Synthases and Xanthocillin Biosynthesis in Aspergillus fumigatus
Abstract
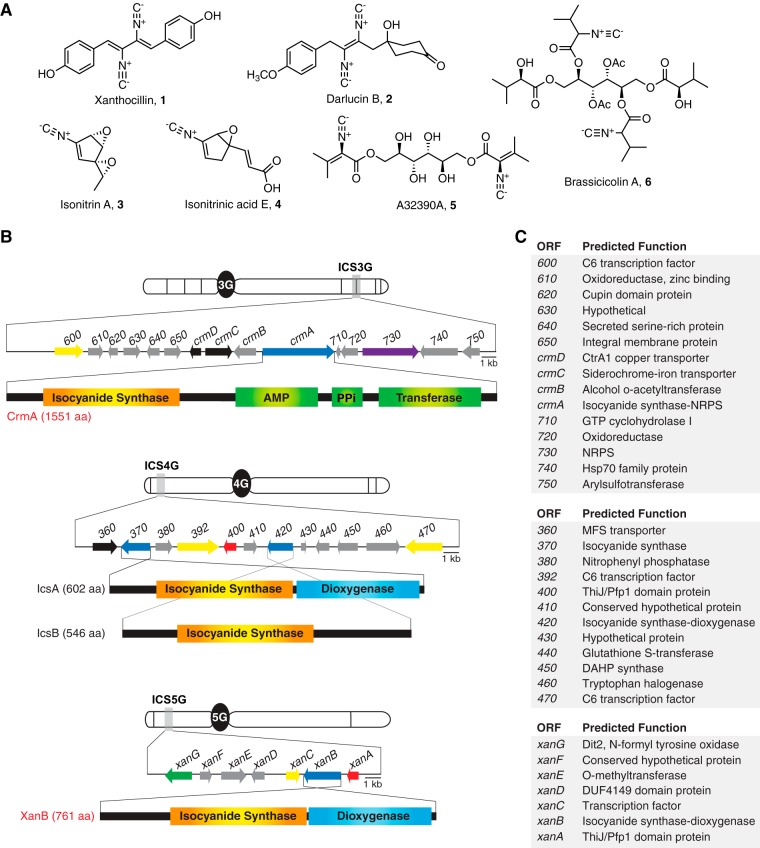
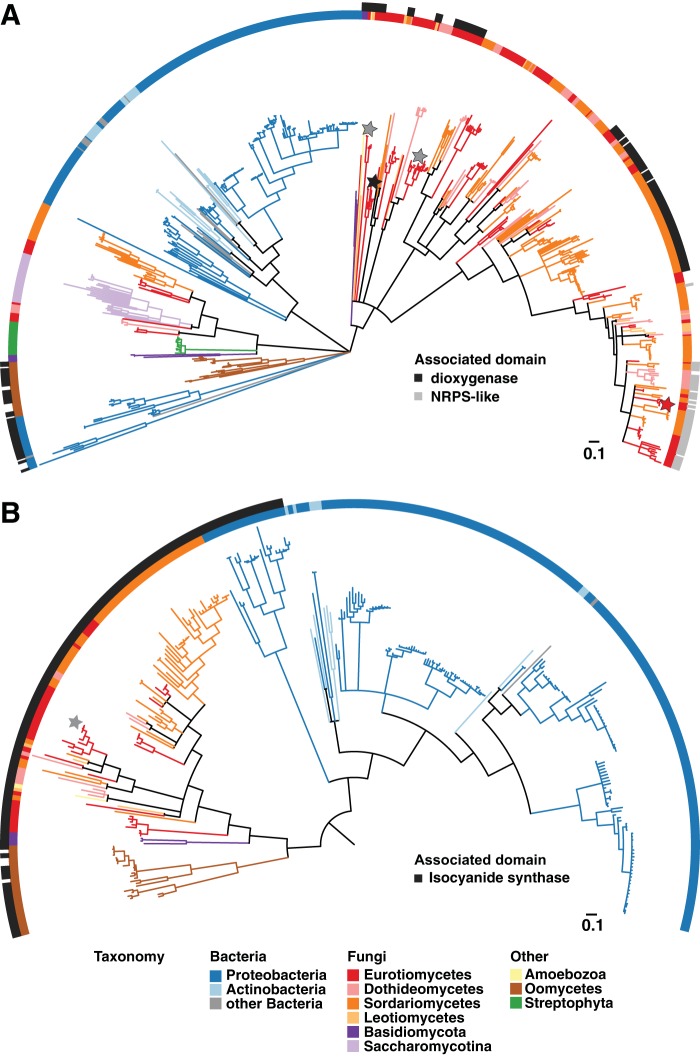
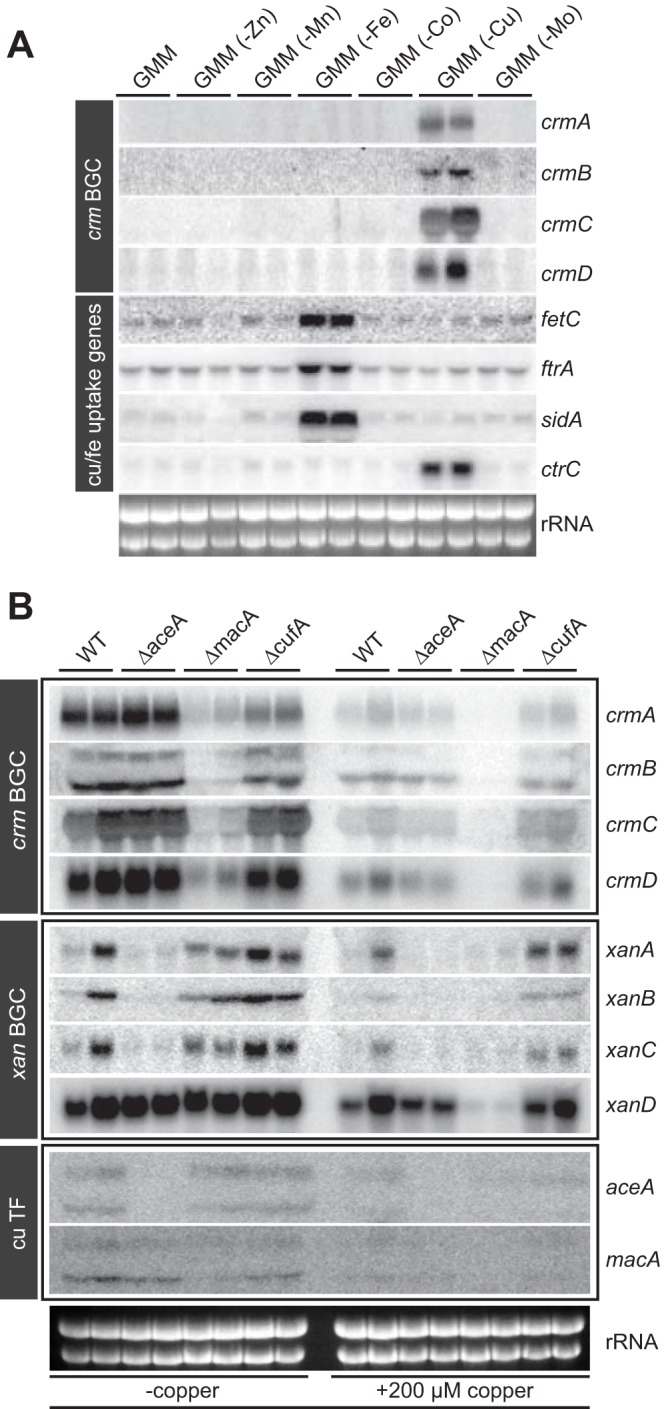
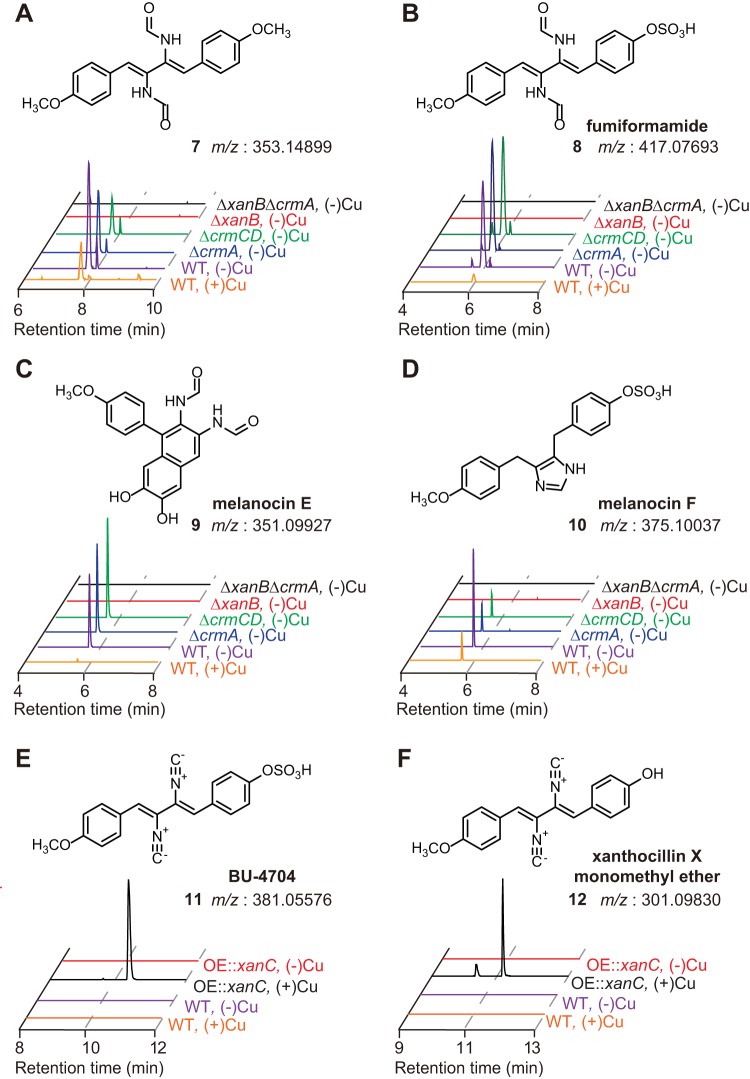
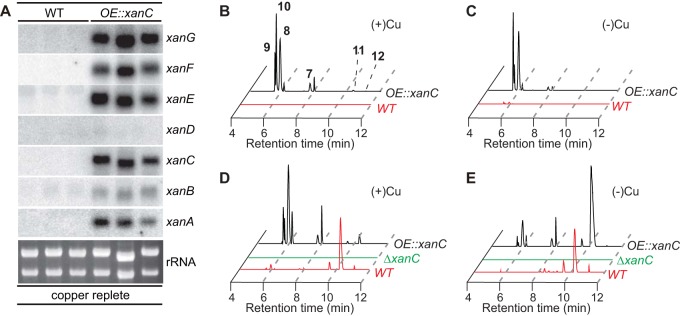
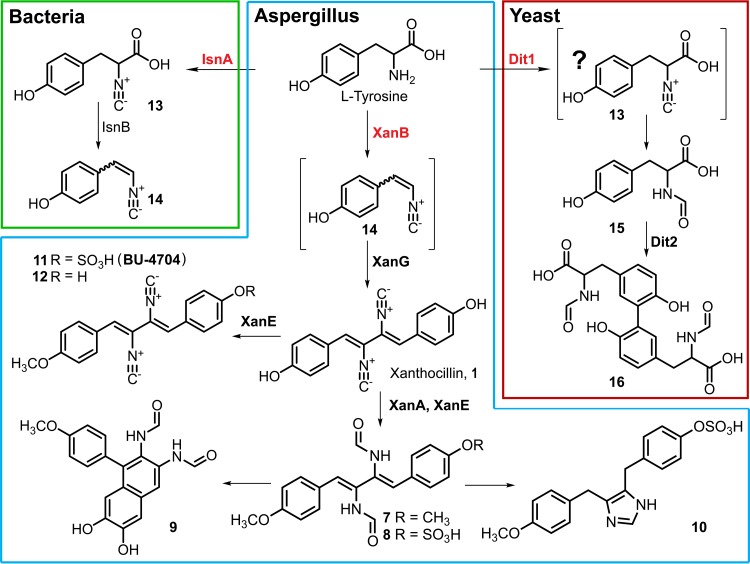
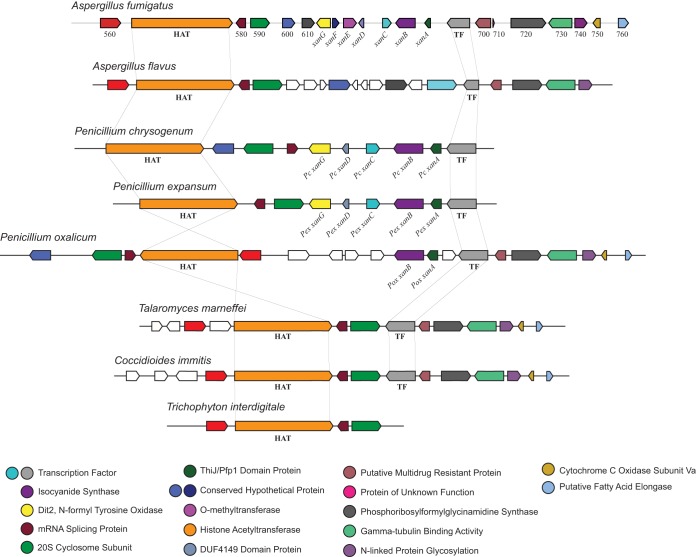
Microbial secondary metabolites, including isocyanide moieties, have been extensively mined for their repertoire of bioactive properties. Although the first naturally occurring isocyanide (xanthocillin) was isolated from the fungus Penicillium notatum over half a century ago, the biosynthetic origins of fungal isocyanides remain unknown. Here we report the identification of a family of isocyanide synthases (ICSs) from the opportunistic human pathogen Aspergillus fumigatus Comparative metabolomics of overexpression or knockout mutants of ICS candidate genes led to the discovery of a fungal biosynthetic gene cluster (BGC) that produces xanthocillin (xan). Detailed analysis of xanthocillin biosynthesis in A. fumigatus revealed several previously undescribed compounds produced by the xan BGC, including two novel members of the melanocin family of compounds. We found both the xan BGC and a second ICS-containing cluster, named the copper-responsive metabolite (crm) BGC, to be transcriptionally responsive to external copper levels and further demonstrated that production of metabolites from the xan BGC is increased during copper starvation. The crm BGC includes a novel type of fungus-specific ICS-nonribosomal peptide synthase (NRPS) hybrid enzyme, CrmA. This family of ICS-NRPS hybrid enzymes is highly enriched in fungal pathogens of humans, insects, and plants. Phylogenetic assessment of all ICSs spanning the tree of life shows not only high prevalence throughout the fungal kingdom but also distribution in species not previously known to harbor BGCs, indicating an untapped resource of fungal secondary metabolism.IMPORTANCE Fungal ICSs are an untapped resource in fungal natural product research. Their isocyanide products have been implicated in plant and insect pathogenesis due to their ability to coordinate transition metals and disable host metalloenzymes. The discovery of a novel isocyanide-producing family of hybrid ICS-NRPS enzymes enriched in medically and agriculturally important fungal pathogens may reveal mechanisms underlying pathogenicity and afford opportunities to discover additional families of isocyanides. Furthermore, the identification of noncanonical ICS BGCs will enable refinement of BGC prediction algorithms to expand on the secondary metabolic potential of fungal and bacterial species. The identification of genes related to ICS BGCs in fungal species not previously known for secondary metabolite-producing capabilities (e.g., Saccharomyces spp.) contributes to our understanding of the evolution of BGC in fungi.
Keywords: Aspergillus fumigatus; copper; isocyanide synthase; melanocin; xanthocillin.
Copyright © 2018 Lim et al.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
