

Smc5/6 Antagonism by HBx Is an Evolutionarily Conserved Function of Hepatitis B Virus Infection in Mammals
- PMID: 29848586
- PMCID: PMC6069175
- DOI: 10.1128/JVI.00769-18
Smc5/6 Antagonism by HBx Is an Evolutionarily Conserved Function of Hepatitis B Virus Infection in Mammals
Abstract
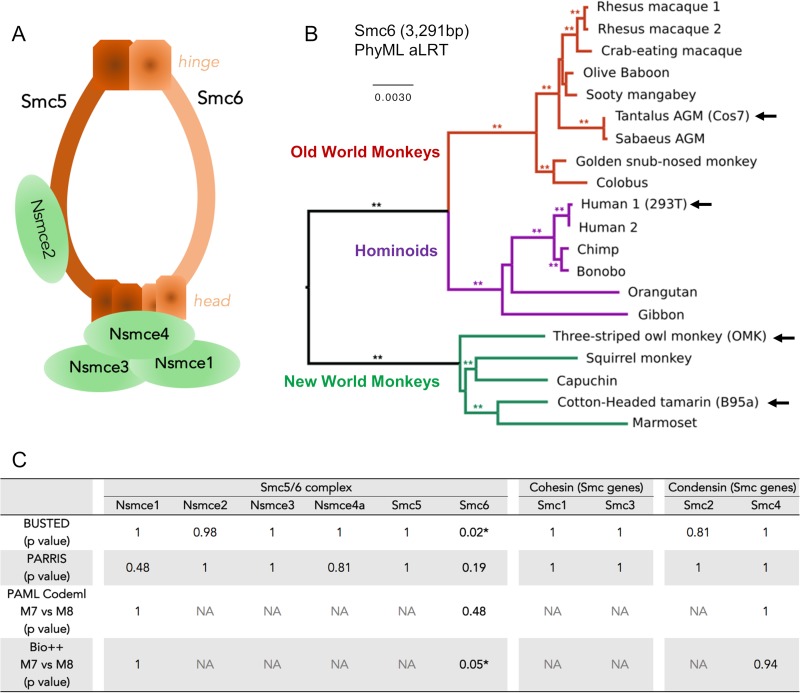
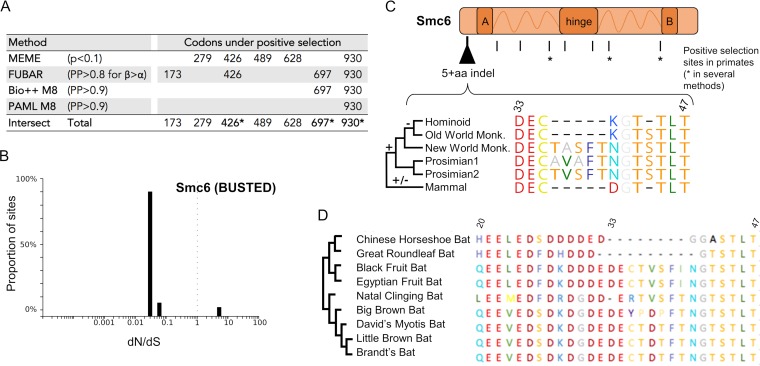
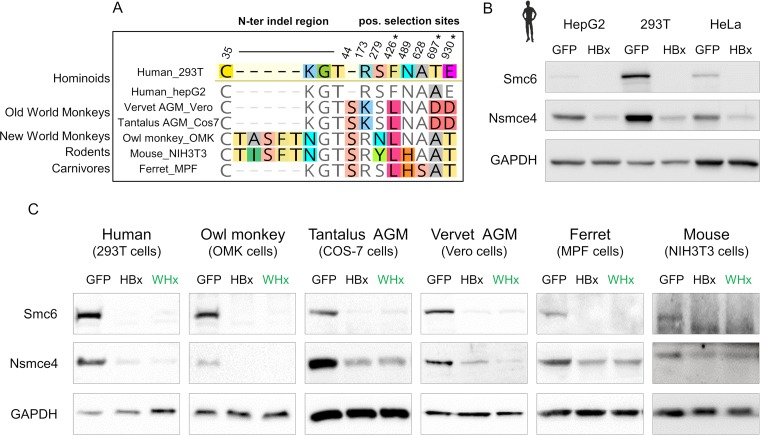
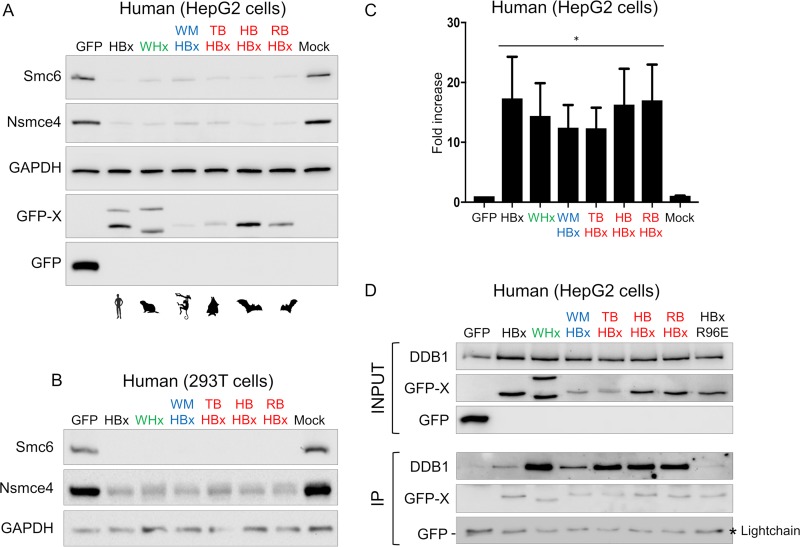
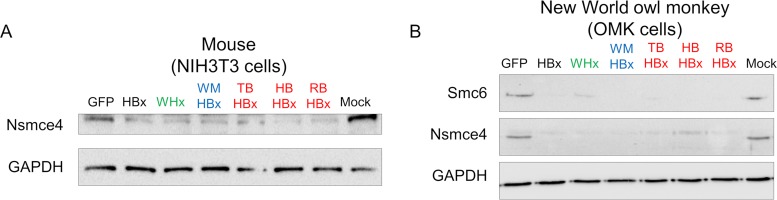
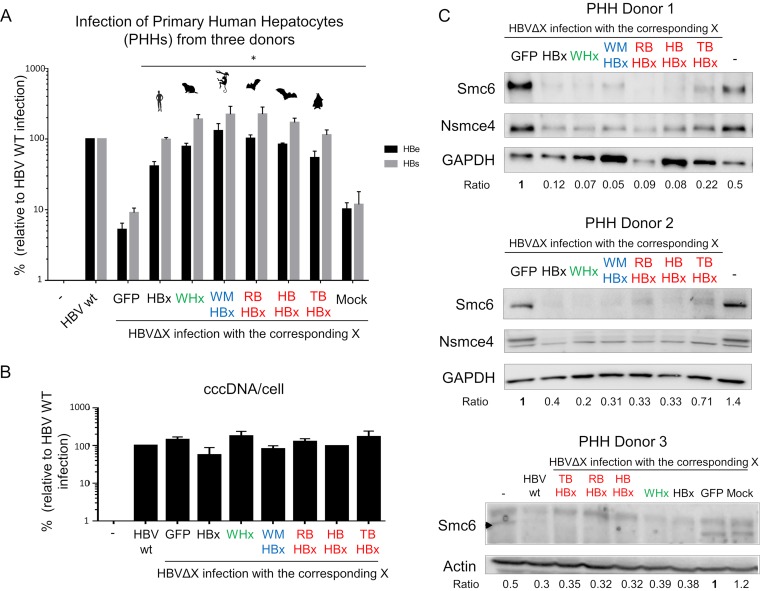
Chronic infection with hepatitis B virus (HBV) is a major cause of liver disease and cancer in humans. HBVs (family Hepadnaviridae) have been associated with mammals for millions of years. Recently, the Smc5/6 complex, known for its essential housekeeping functions in genome maintenance, was identified as an antiviral restriction factor of human HBV. The virus has, however, evolved to counteract this defense mechanism by degrading the complex via its regulatory HBx protein. Whether the antiviral activity of the Smc5/6 complex against hepadnaviruses is an important and evolutionarily conserved function is unknown. In this study, we used an evolutionary and functional approach to address this question. We first performed phylogenetic and positive selection analyses of the Smc5/6 complex subunits and found that they have been conserved in primates and mammals. Yet, Smc6 showed marks of adaptive evolution, potentially reminiscent of a virus-host "arms race." We then functionally tested the HBx proteins from six divergent hepadnaviruses naturally infecting primates, rodents, and bats. We demonstrate that despite little sequence homology, these HBx proteins efficiently degraded mammalian Smc5/6 complexes, independently of the host species and of the sites under positive selection. Importantly, all HBx proteins also rescued the replication of an HBx-deficient HBV in primary human hepatocytes. These findings point to an evolutionarily conserved requirement for Smc5/6 inactivation by HBx, showing that Smc5/6 antiviral activity has been an important defense mechanism against hepadnaviruses in mammals. It will be interesting to investigate whether Smc5/6 may further be a restriction factor of other, yet-unidentified viruses that may have driven some of its adaptation.IMPORTANCE Infection with hepatitis B virus (HBV) led to 887,000 human deaths in 2015. HBV has been coevolving with mammals for millions of years. Recently, the Smc5/6 complex, which has essential housekeeping functions, was identified as a restriction factor of human HBV antagonized by the regulatory HBx protein. Here we address whether the antiviral activity of Smc5/6 is an important evolutionarily conserved function. We found that all six subunits of Smc5/6 have been conserved in primates, with only Smc6 showing signatures of an "evolutionary arms race." Using evolution-guided functional analyses that included infections of primary human hepatocytes, we demonstrated that HBx proteins from very divergent mammalian HBVs could all efficiently antagonize Smc5/6, independently of the host species and sites under positive selection. These findings show that Smc5/6 antiviral activity against HBV is an important function in mammals. They also raise the intriguing possibility that Smc5/6 may restrict other, yet-unidentified viruses.
Keywords: HBx; Smc5/6 complex; antagonism; evolution of virus and host genes; hepatitis B virus; positive selection; restriction factor; virus-host interaction.
Copyright © 2018 American Society for Microbiology.
Figures

References
-
- Lauber C, Seitz S, Mattei S, Suh A, Beck J, Herstein J, Borold J, Salzburger W, Kaderali L, Briggs JAG, Bartenschlager R. 2017. Deciphering the origin and evolution of hepatitis B viruses by means of a family of non-enveloped fish viruses. Cell Host Microbe 22:387–399.e386. doi: 10.1016/j.chom.2017.07.019. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
