

A Suppressor Screen for AGO1 Degradation by the Viral F-Box P0 Protein Uncovers a Role for AGO DUF1785 in sRNA Duplex Unwinding
- PMID: 29848768
- PMCID: PMC6048793
- DOI: 10.1105/tpc.18.00111
A Suppressor Screen for AGO1 Degradation by the Viral F-Box P0 Protein Uncovers a Role for AGO DUF1785 in sRNA Duplex Unwinding
Abstract
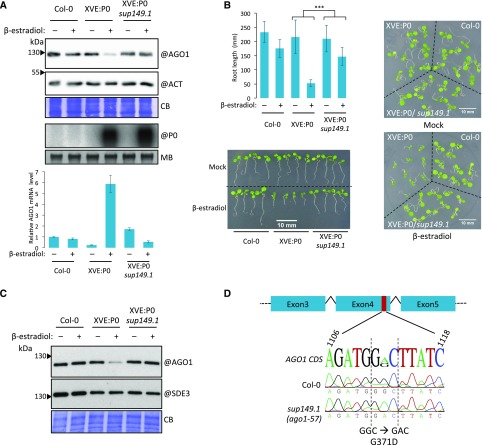
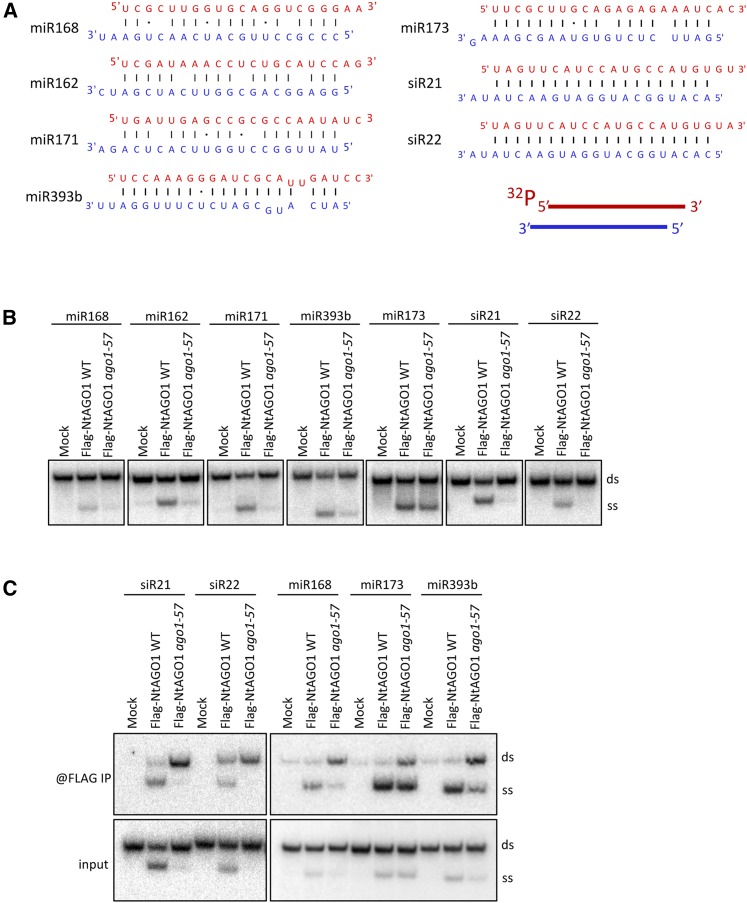
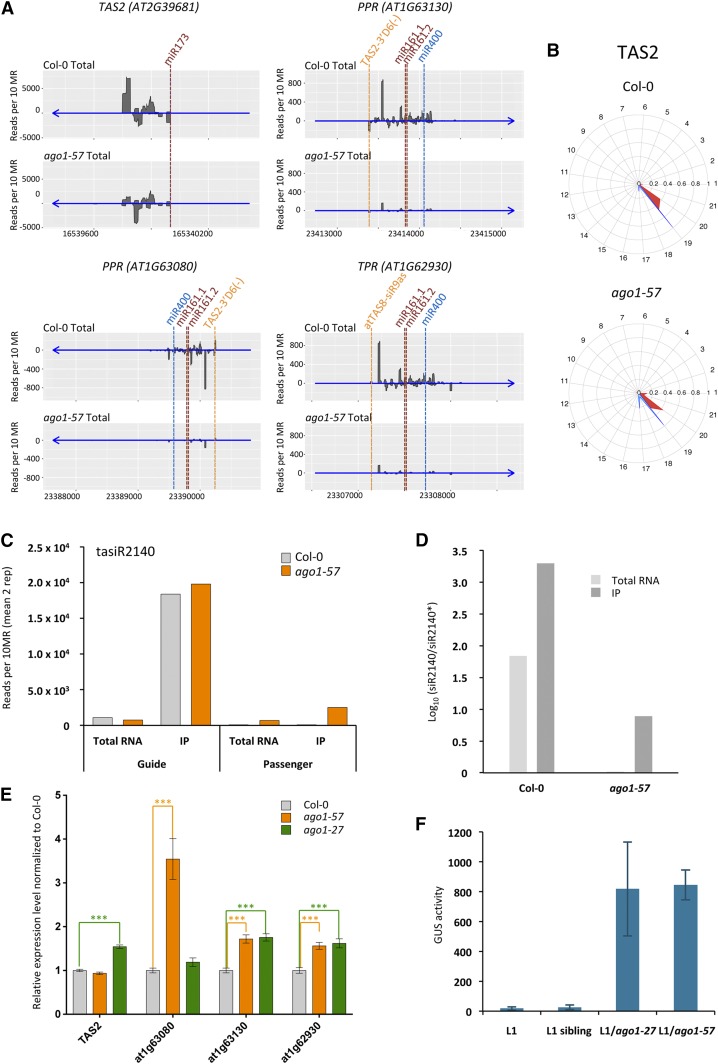
In Arabidopsis thaliana, ARGONAUTE1 (AGO1) plays a central role in microRNA (miRNA) and small interfering RNA (siRNA)-mediated silencing and is a key component in antiviral responses. The polerovirus F-box P0 protein triggers AGO1 degradation as a viral counterdefense. Here, we identified a motif in AGO1 that is required for its interaction with the S phase kinase-associated protein1-cullin 1-F-box protein (SCF) P0 (SCFP0) complex and subsequent degradation. The AGO1 P0 degron is conserved and confers P0-mediated degradation to other AGO proteins. Interestingly, the degron motif is localized in the DUF1785 domain of AGO1, in which a single point mutation (ago1-57, obtained by forward genetic screening) compromises recognition by SCFP0 Recapitulating formation of the RNA-induced silencing complex in a cell-free system revealed that this mutation impairs RNA unwinding, leading to stalled forms of AGO1 still bound to double-stranded RNAs. In vivo, the DUF1785 is required for unwinding perfectly matched siRNA duplexes, but is mostly dispensable for unwinding imperfectly matched miRNA duplexes. Consequently, its mutation nearly abolishes phased siRNA production and sense transgene posttranscriptional gene silencing. Overall, our work sheds new light on the mode of AGO1 recognition by P0 and the in vivo function of DUF1785 in RNA silencing.
© 2018 American Society of Plant Biologists. All rights reserved.
Figures





References
-
- Almasi R., Miller W.A., Ziegler-Graff V. (2015). Mild and severe cereal yellow dwarf viruses differ in silencing suppressor efficiency of the P0 protein. Virus Res. 208: 199–206. - PubMed
-
- Arribas-Hernández L., Marchais A., Poulsen C., Haase B., Hauptmann J., Benes V., Meister G., Brodersen P. (2016b). The slicer activity of ARGONAUTE1 is required specifically for the phasing, not production, of trans-acting short interfering RNAs in Arabidopsis. Plant Cell 28: 1563–1580. - PMC - PubMed
-
- Baumberger N., Tsai C.H., Lie M., Havecker E., Baulcombe D.C. (2007). The Polerovirus silencing suppressor P0 targets ARGONAUTE proteins for degradation. Curr. Biol. 17: 1609–1614. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
