

Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells
- PMID: 29849141
- PMCID: PMC6320248
- DOI: 10.1038/s41586-018-0178-z
Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells
Abstract
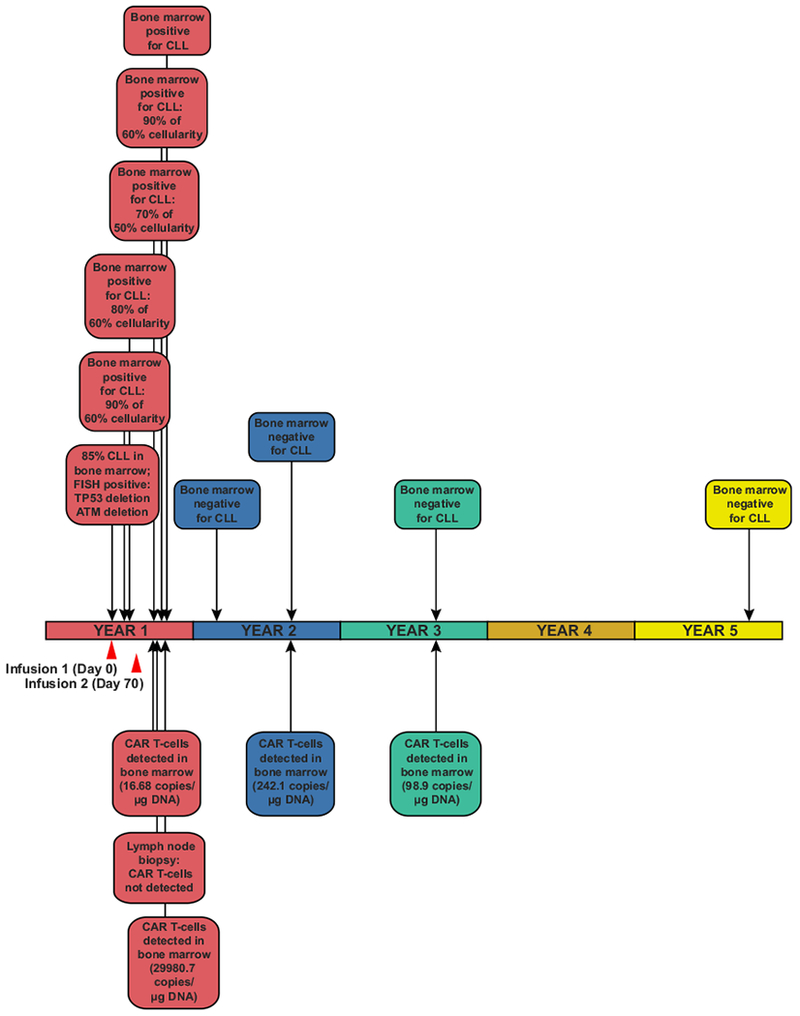
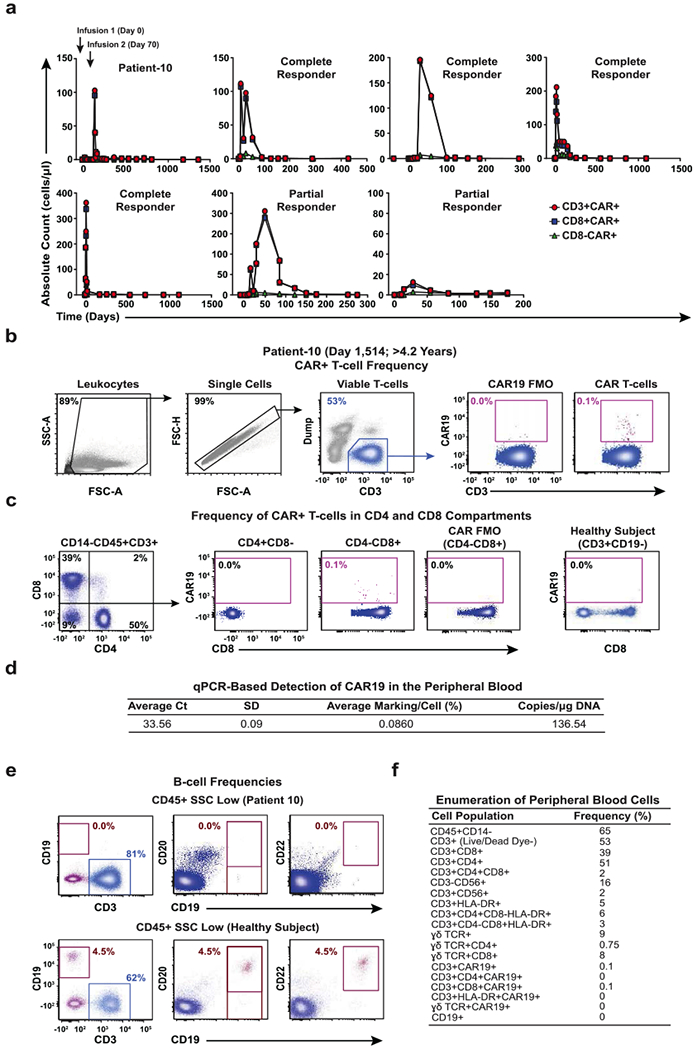
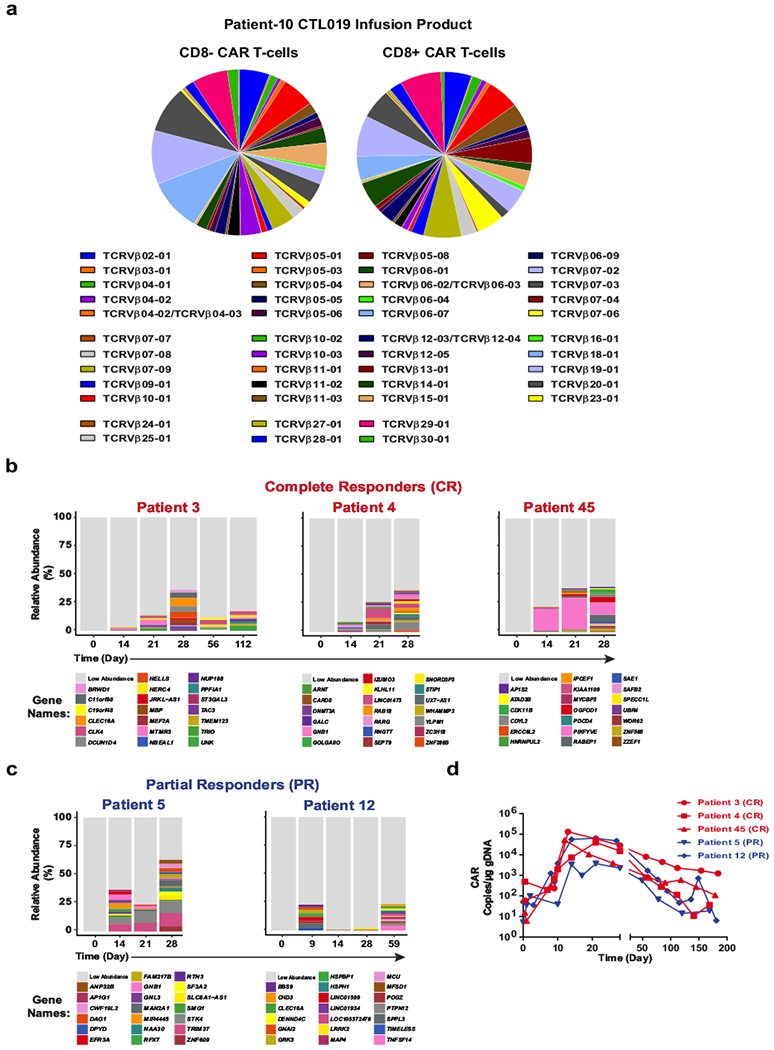
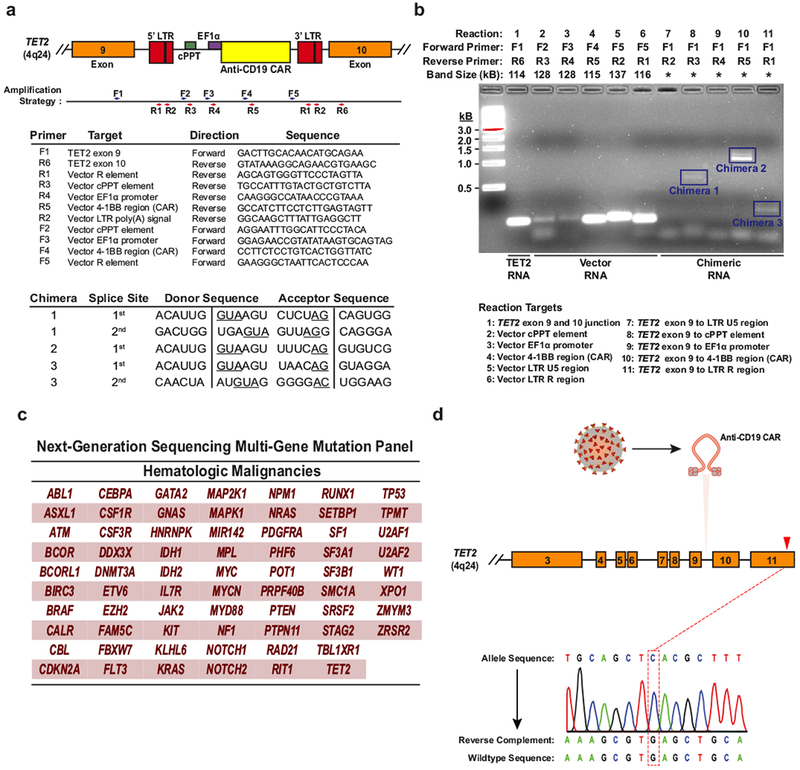
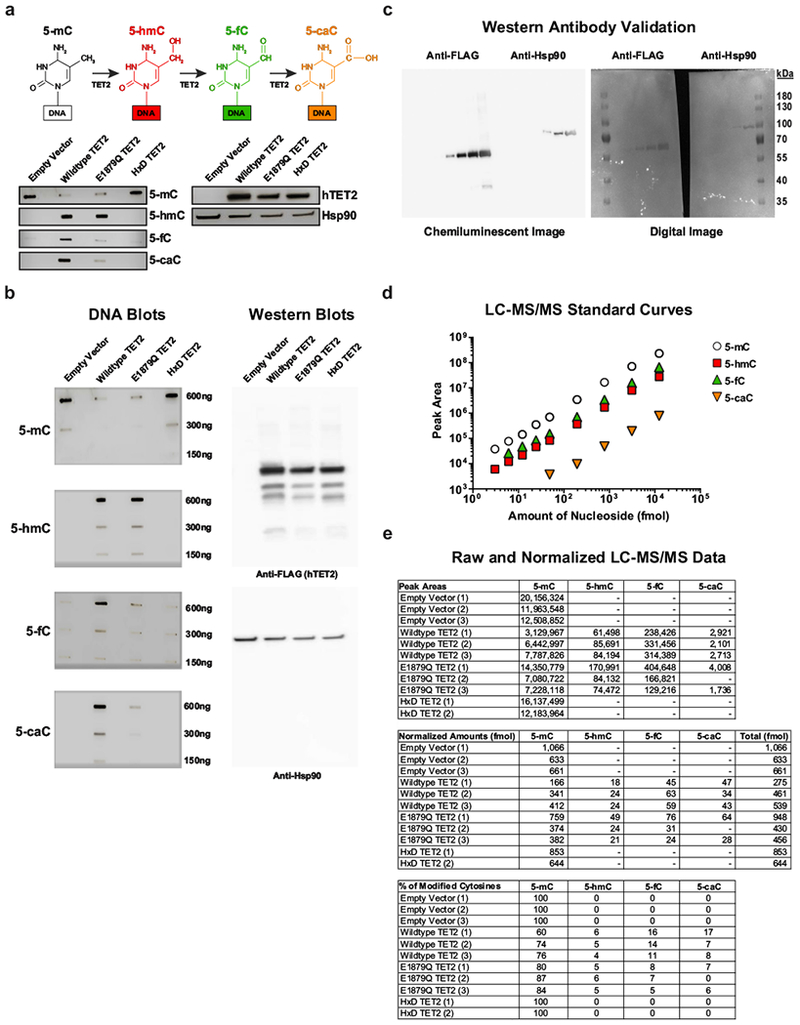
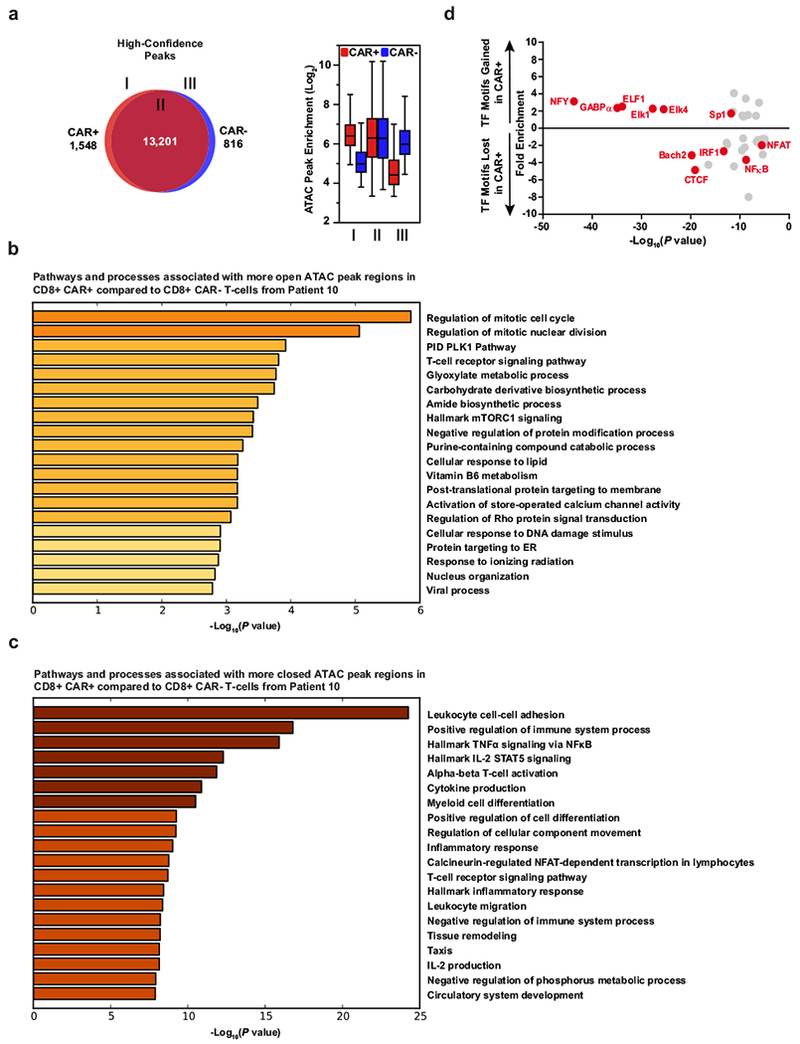
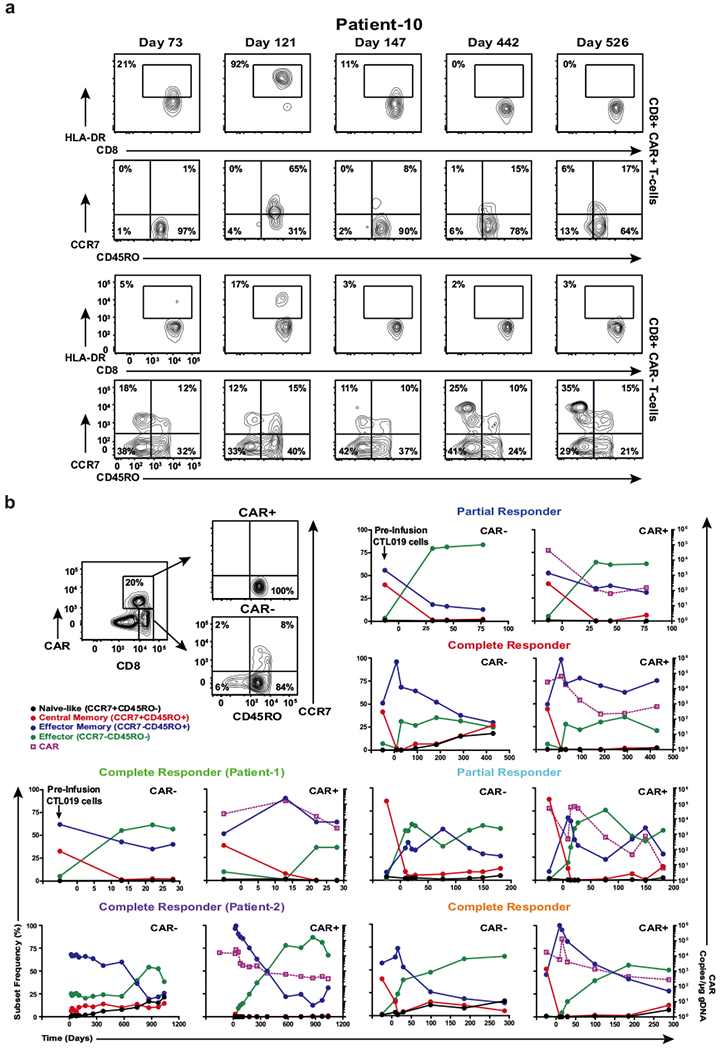
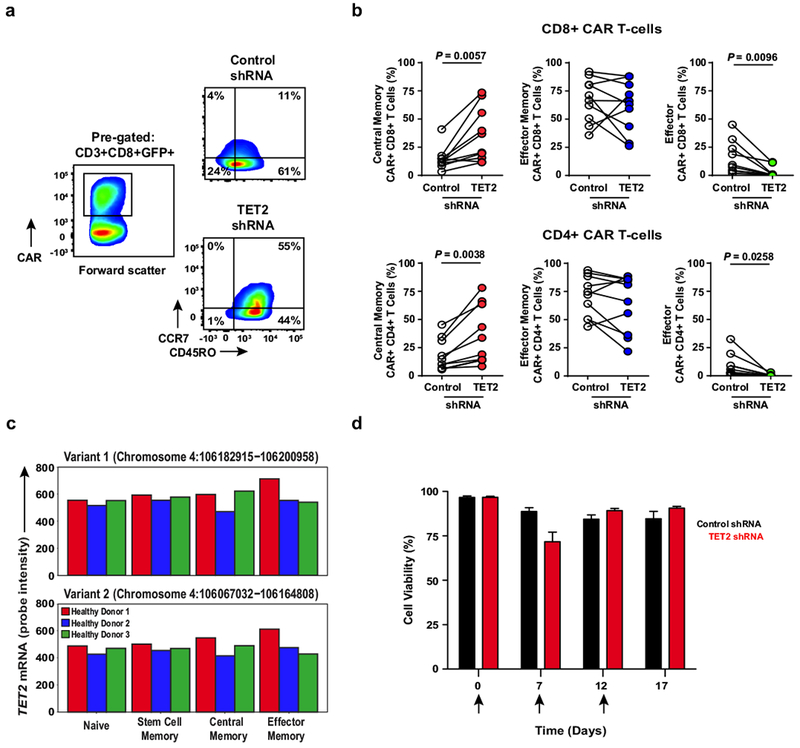
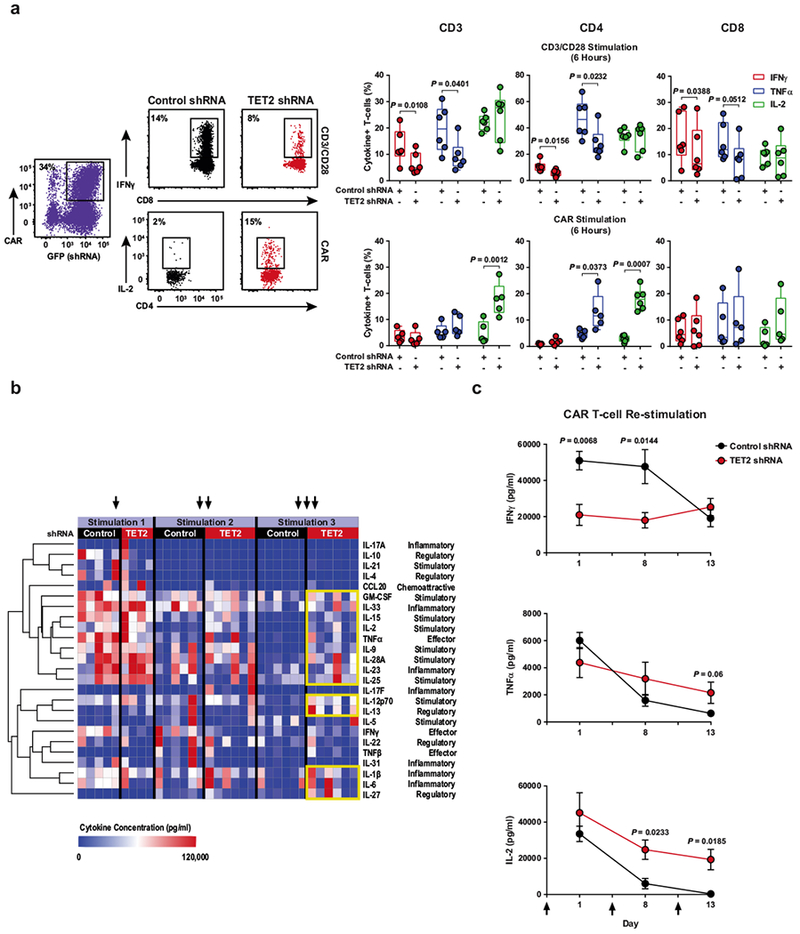
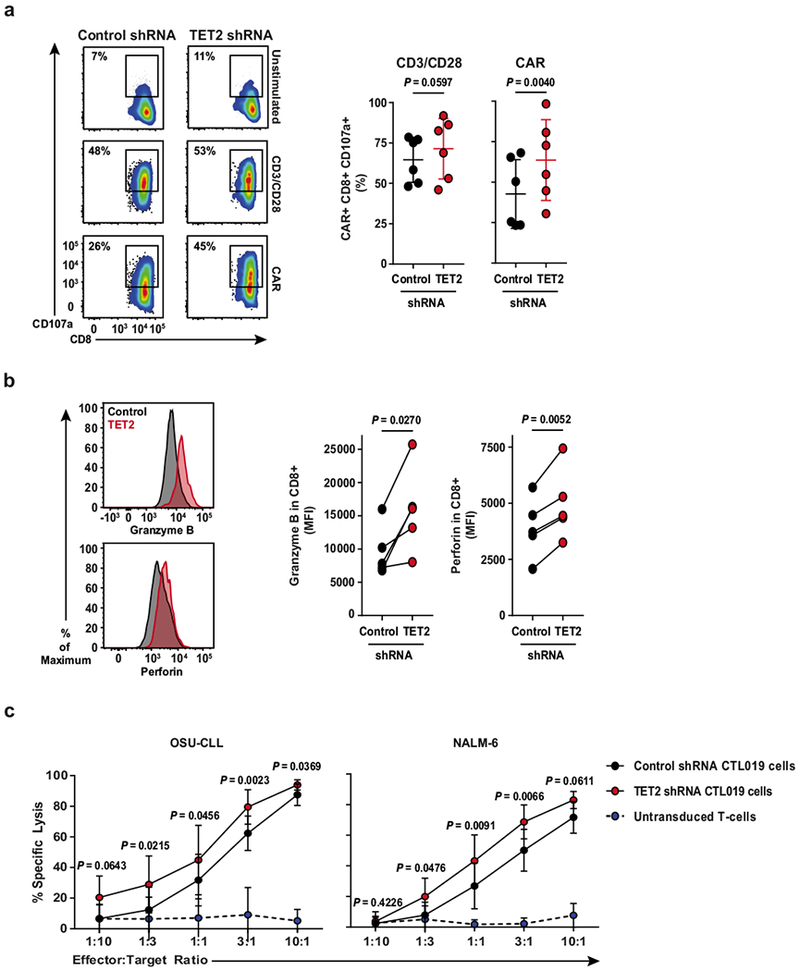
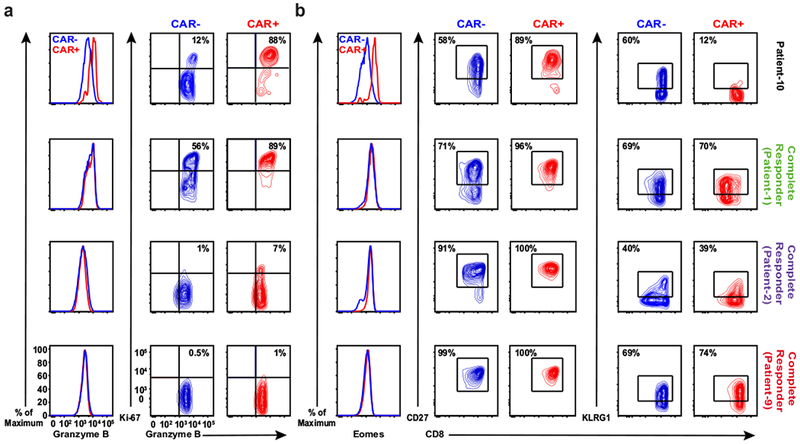
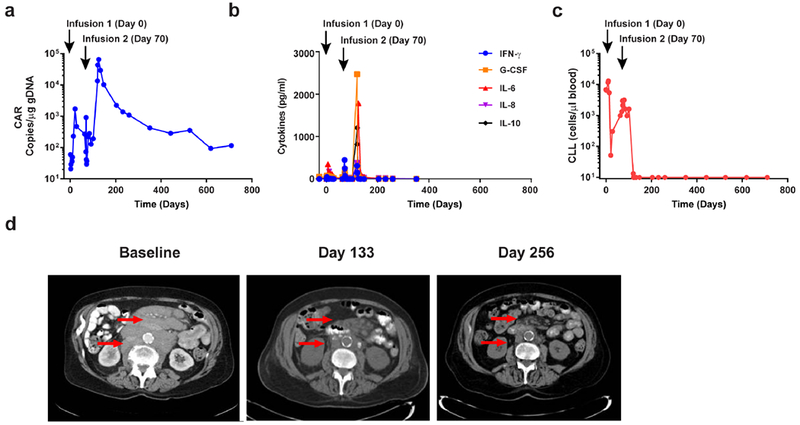
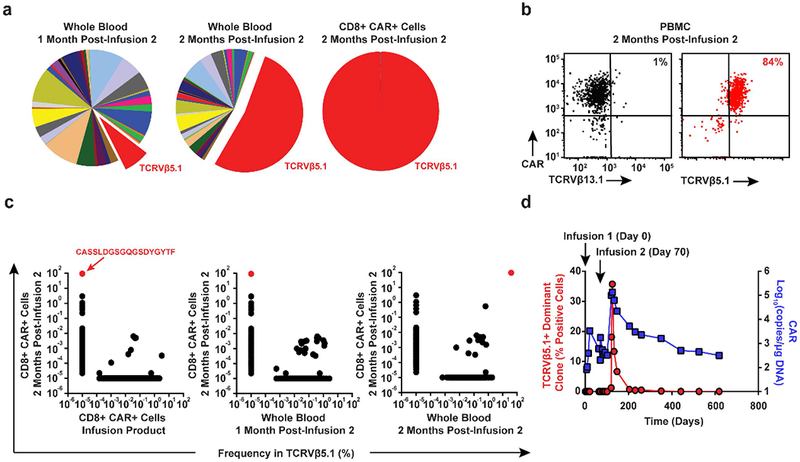
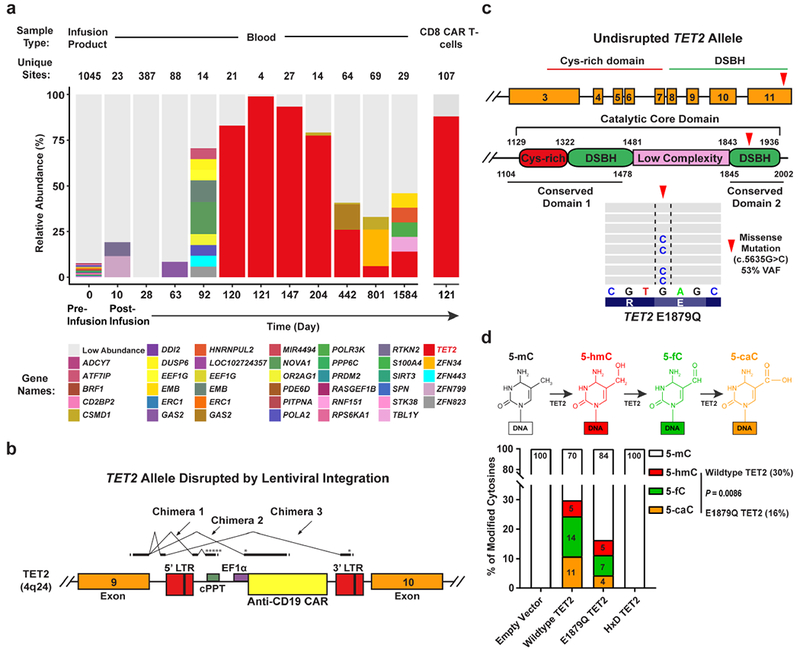
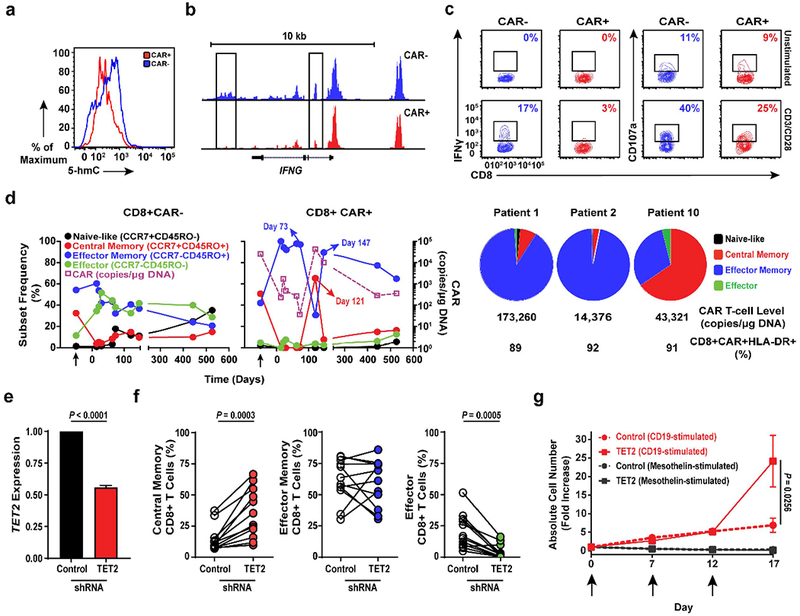
Cancer immunotherapy based on genetically redirecting T cells has been used successfully to treat B cell malignancies1-3. In this strategy, the T cell genome is modified by integration of viral vectors or transposons encoding chimaeric antigen receptors (CARs) that direct tumour cell killing. However, this approach is often limited by the extent of expansion and persistence of CAR T cells4,5. Here we report mechanistic insights from studies of a patient with chronic lymphocytic leukaemia treated with CAR T cells targeting the CD19 protein. Following infusion of CAR T cells, anti-tumour activity was evident in the peripheral blood, lymph nodes and bone marrow; this activity was accompanied by complete remission. Unexpectedly, at the peak of the response, 94% of CAR T cells originated from a single clone in which lentiviral vector-mediated insertion of the CAR transgene disrupted the methylcytosine dioxygenase TET2 gene. Further analysis revealed a hypomorphic mutation in this patient's second TET2 allele. TET2-disrupted CAR T cells exhibited an epigenetic profile consistent with altered T cell differentiation and, at the peak of expansion, displayed a central memory phenotype. Experimental knockdown of TET2 recapitulated the potency-enhancing effect of TET2 dysfunction in this patient's CAR T cells. These findings suggest that the progeny of a single CAR T cell induced leukaemia remission and that TET2 modification may be useful for improving immunotherapies.
Conflict of interest statement
Competing Financial Interests
J.A.F., C.L.N., R.M.Y., B.L.L. M.K., D.L.P., S.F.L., F.D.B., C.H.J. and J.J.M. hold patents related to CTL019 cell therapy. These authors declare no additional financial interests. The remaining authors declare no competing financial interests.
Figures
Comment in
-
Tumour tamed by transfer of one T cell.Nature. 2018 Jun;558(7709):193-195. doi: 10.1038/d41586-018-05251-5. Nature. 2018. PMID: 29895908 No abstract available.
-
Success stories.Nat Rev Cancer. 2018 Aug;18(8):465. doi: 10.1038/s41568-018-0039-y. Nat Rev Cancer. 2018. PMID: 29934503 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
