

Role of oxytocin in the ventral tegmental area in social reinforcement
- PMID: 29852406
- PMCID: PMC6109598
- DOI: 10.1016/j.psyneuen.2018.05.028
Role of oxytocin in the ventral tegmental area in social reinforcement
Abstract
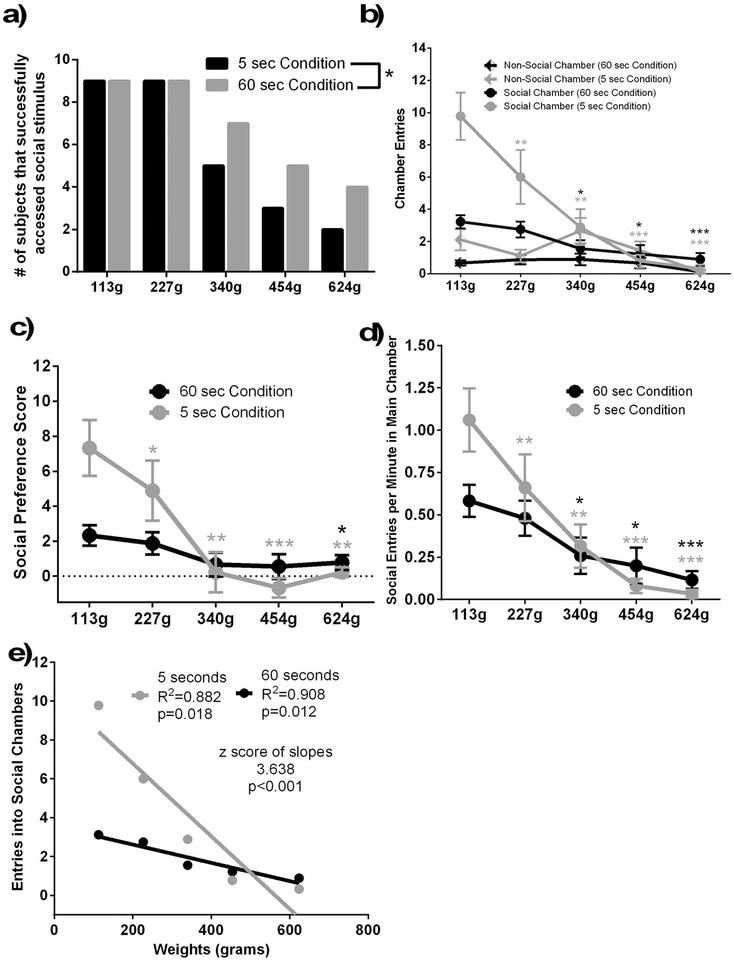
The rewarding properties of social interactions play a critical role in the development and maintenance of social relationships, and deficits in social reward are associated with various psychiatric disorders. In the present study, we used a novel Operant Social Preference (OSP) task to investigate the reinforcing properties of social interactions under conditions of high or low reward value, and high or low behavioral effort in male Syrian hamsters. Further, we investigated the role of oxytocin (OT) in a key structure of the mesolimbic reward system, the ventral tegmental area (VTA), in mediating the reinforcing properties of social interaction. Adult male hamsters were placed in a three-chambered apparatus, and allowed access to either a social chamber containing an unrestrained conspecific or a non-social chamber, by pushing through a one-way entry, vertical-swing door. Increasing the duration of social interaction (reward value) decreased the frequency of entering the social interaction chambers, whereas decreasing the duration of social interaction conversely increased the frequency of entries. Moreover, increasing behavioral effort required to access social interaction decreased the frequency of entries, especially under conditions when the duration of social interaction was only 5 s. OT injected into the VTA decreased the frequency of entering social interaction chambers in a manner similar to that observed when duration was increased, whereas injection of an OT receptor antagonist in the VTA increased the frequency of seeking social interaction. Taken together, these data support the hypothesis that activation of OT receptors in the VTA are critical for the reinforcing properties of social interactions. Furthermore, social interactions may exhibit duration and cost dependent reinforcing effects on behavior similar to those observed with food and drugs of abuse.
Keywords: Dopamine; Mesolimbic dopamine system; Neuropeptides; Operant; Social behavior; Social cognition; Social interaction; Social motivation; Social reward; Social salience; Vasopressin.
Copyright © 2018 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare no conflicts of interest.
The authors have no competing interests to declare.
Figures







References
-
- Albers HE, 2012. The regulation of social recognition, social communication and aggression: vasopressin in the social behavior neural network. Horm. Behav 61, 283–292. - PubMed
-
- Angioni L, Cocco C, Ferri GL, Argiolas A, Melis MR, Sanna F, 2016. Involvement of nigral oxytocin in locomotor activity: a behavioral, immunohistochemical and lesion study in male rats. Horm. Behav 83, 23–38. - PubMed
-
- Bardo MT, Bevins RA, 2000. Conditioned place preference: what does it add to our preclinical understanding of drug reward? Psychopharmacology 153 (3), 1–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
