

Glial βII Spectrin Contributes to Paranode Formation and Maintenance
- PMID: 29853631
- PMCID: PMC6031582
- DOI: 10.1523/JNEUROSCI.3647-17.2018
Glial βII Spectrin Contributes to Paranode Formation and Maintenance
Abstract
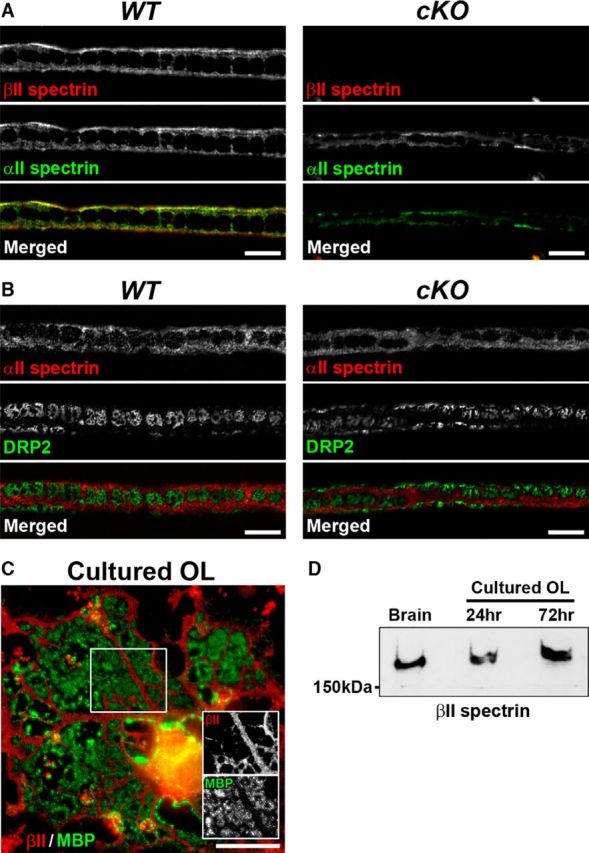
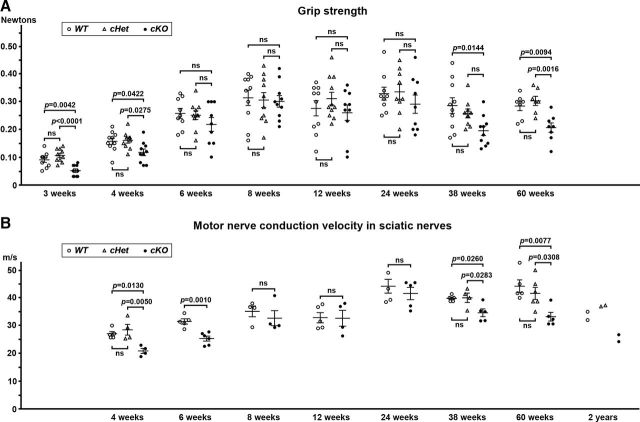
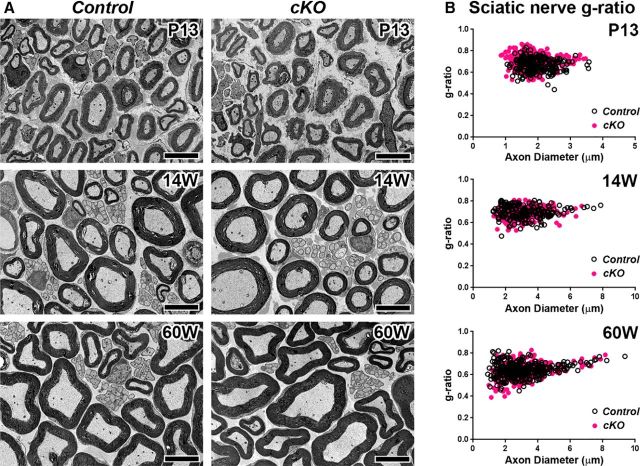
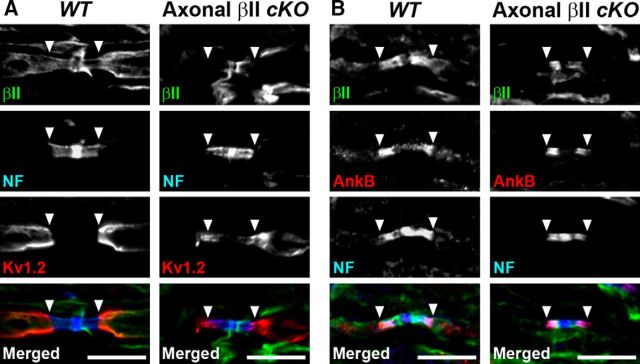
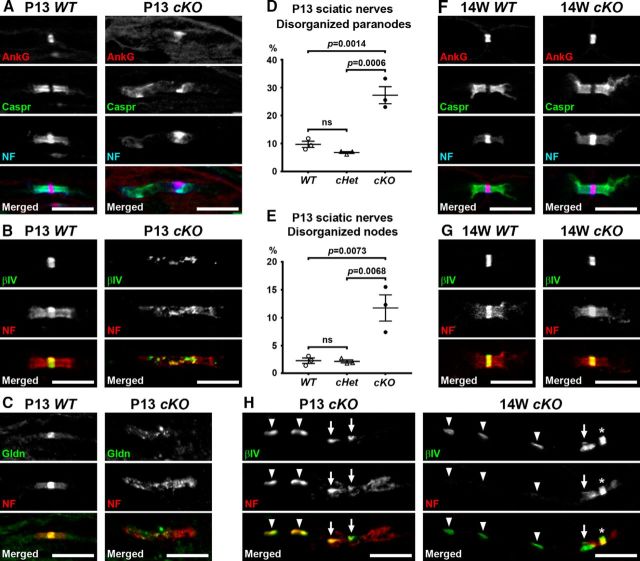
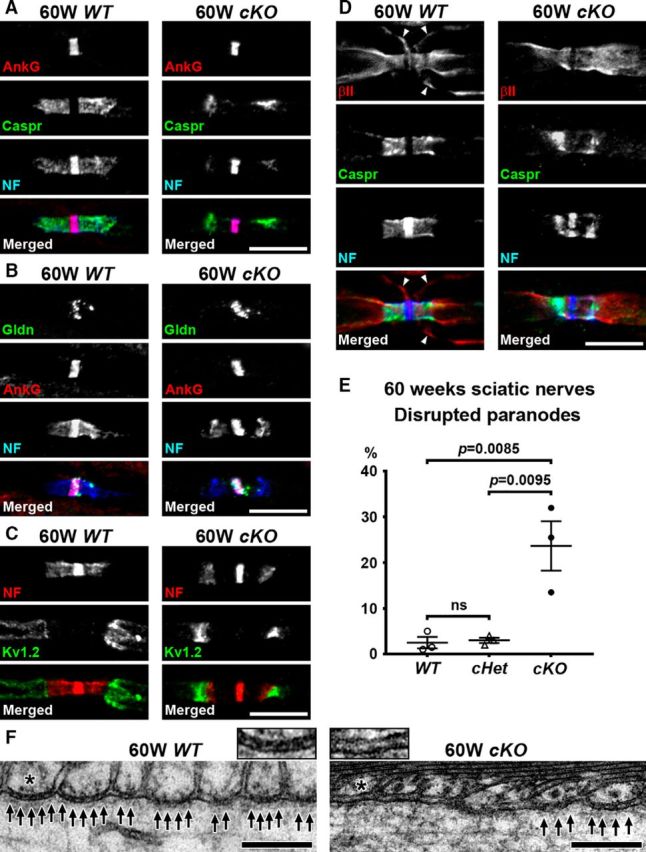
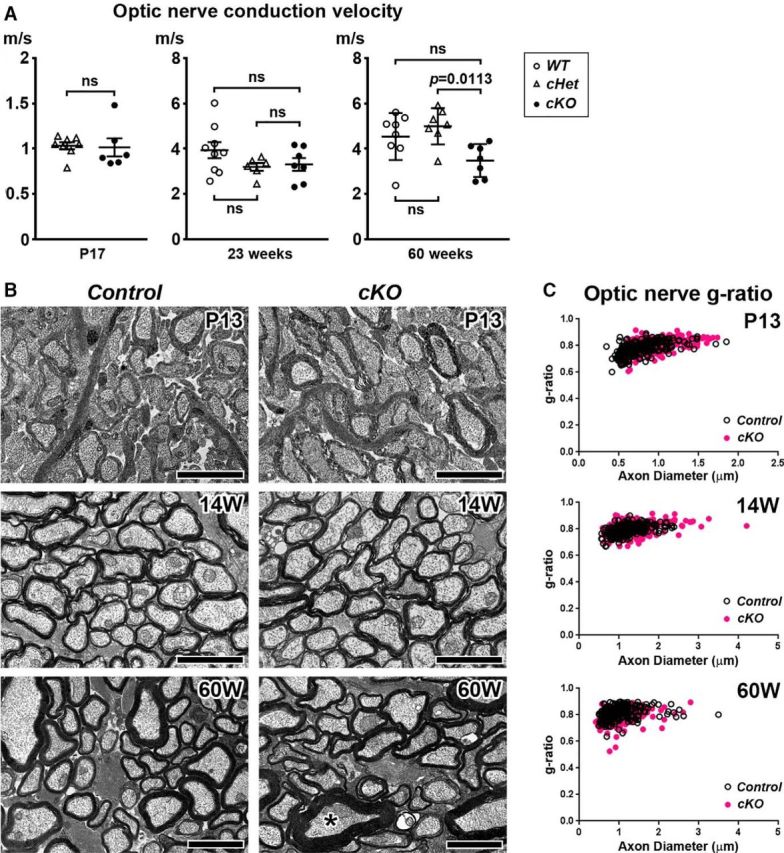
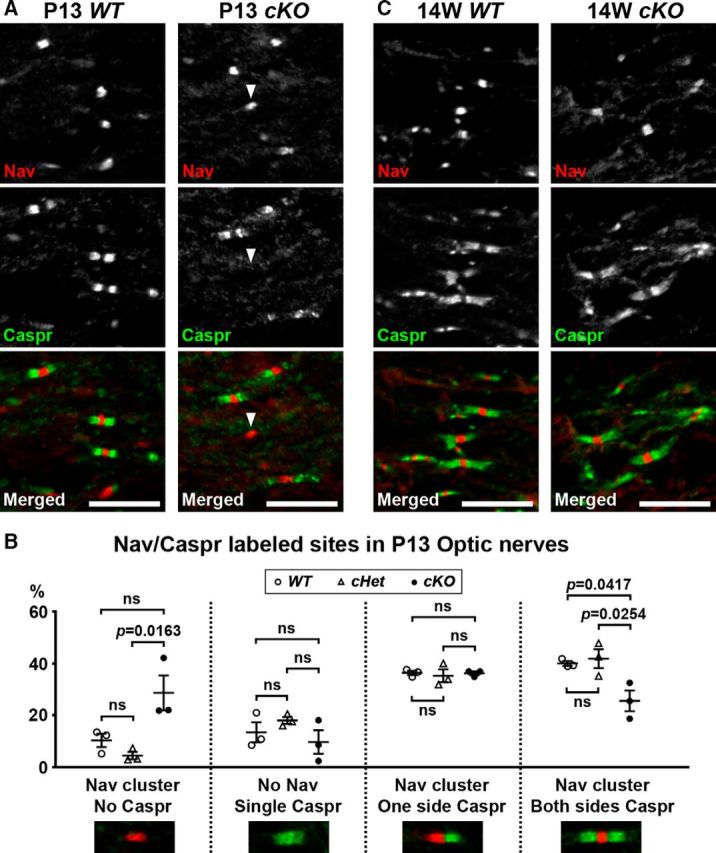
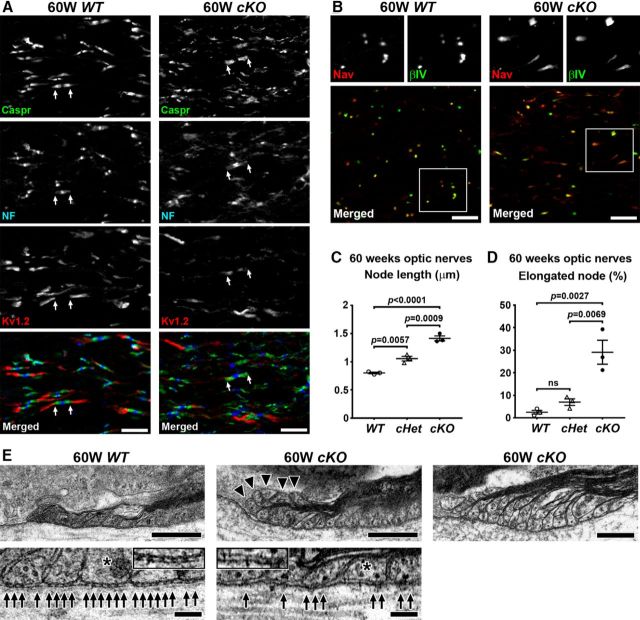
Action potential conduction along myelinated axons depends on high densities of voltage-gated Na+ channels at the nodes of Ranvier. Flanking each node, paranodal junctions (paranodes) are formed between axons and Schwann cells in the peripheral nervous system (PNS) or oligodendrocytes in the CNS. Paranodal junctions contribute to both node assembly and maintenance. Despite their importance, the molecular mechanisms responsible for paranode assembly and maintenance remain poorly understood. βII spectrin is expressed in diverse cells and is an essential part of the submembranous cytoskeleton. Here, we show that Schwann cell βII spectrin is highly enriched at paranodes. To elucidate the roles of glial βII spectrin, we generated mutant mice lacking βII spectrin in myelinating glial cells by crossing mice with a floxed allele of Sptbn1 with Cnp-Cre mice, and analyzed both male and female mice. Juvenile (4 weeks) and middle-aged (60 weeks) mutant mice showed reduced grip strength and sciatic nerve conduction slowing, whereas no phenotype was observed between 8 and 24 weeks of age. Consistent with these findings, immunofluorescence microscopy revealed disorganized paranodes in the PNS and CNS of both postnatal day 13 and middle-aged mutant mice, but not in young adult mutant mice. Electron microscopy confirmed partial loss of transverse bands at the paranodal axoglial junction in the middle-aged mutant mice in both the PNS and CNS. These findings demonstrate that a spectrin-based cytoskeleton in myelinating glia contributes to formation and maintenance of paranodal junctions.SIGNIFICANCE STATEMENT Myelinating glia form paranodal axoglial junctions that flank both sides of the nodes of Ranvier. These junctions contribute to node formation and maintenance and are essential for proper nervous system function. We found that a submembranous spectrin cytoskeleton is highly enriched at paranodes in Schwann cells. Ablation of βII spectrin in myelinating glial cells disrupted the paranodal cell adhesion complex in both peripheral and CNSs, resulting in muscle weakness and sciatic nerve conduction slowing in juvenile and middle-aged mice. Our data show that a spectrin-based submembranous cytoskeleton in myelinating glia plays important roles in paranode formation and maintenance.
Keywords: myelin; node of Ranvier; paranode; spectrin.
Copyright © 2018 the authors 0270-6474/18/386063-13$15.00/0.
Figures
References
-
- Bhat MA, Rios JC, Lu Y, Garcia-Fresco GP, Ching W, St Martin M, Li J, Einheber S, Chesler M, Rosenbluth J, Salzer JL, Bellen HJ (2001) Axon-glia interactions and the domain organization of myelinated axons requires neurexin IV/Caspr/Paranodin. Neuron 30:369–383. 10.1016/S0896-6273(01)00294-X - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous