

Non-equilibrium repressor binding kinetics link DNA damage dose to transcriptional timing within the SOS gene network
- PMID: 29856734
- PMCID: PMC5999292
- DOI: 10.1371/journal.pgen.1007405
Non-equilibrium repressor binding kinetics link DNA damage dose to transcriptional timing within the SOS gene network
Abstract
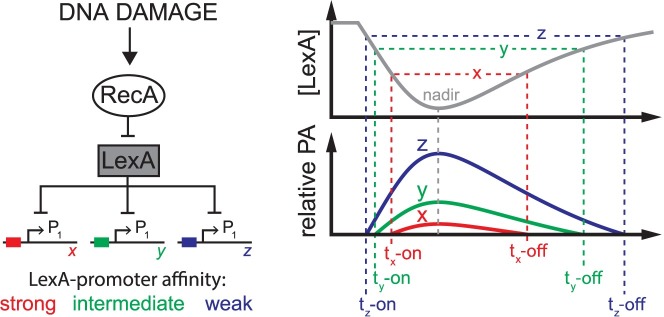
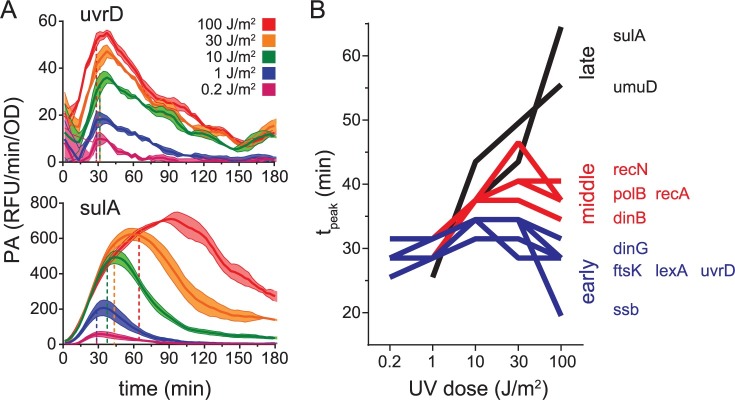
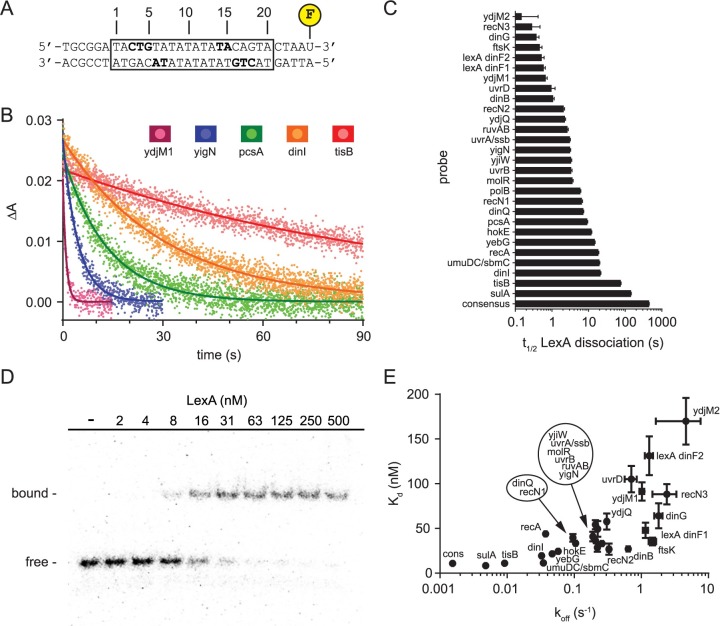
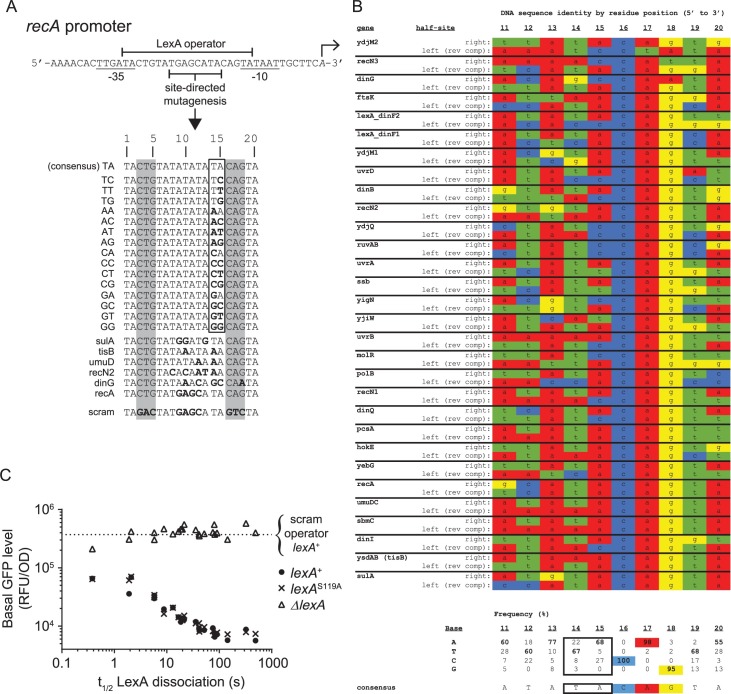
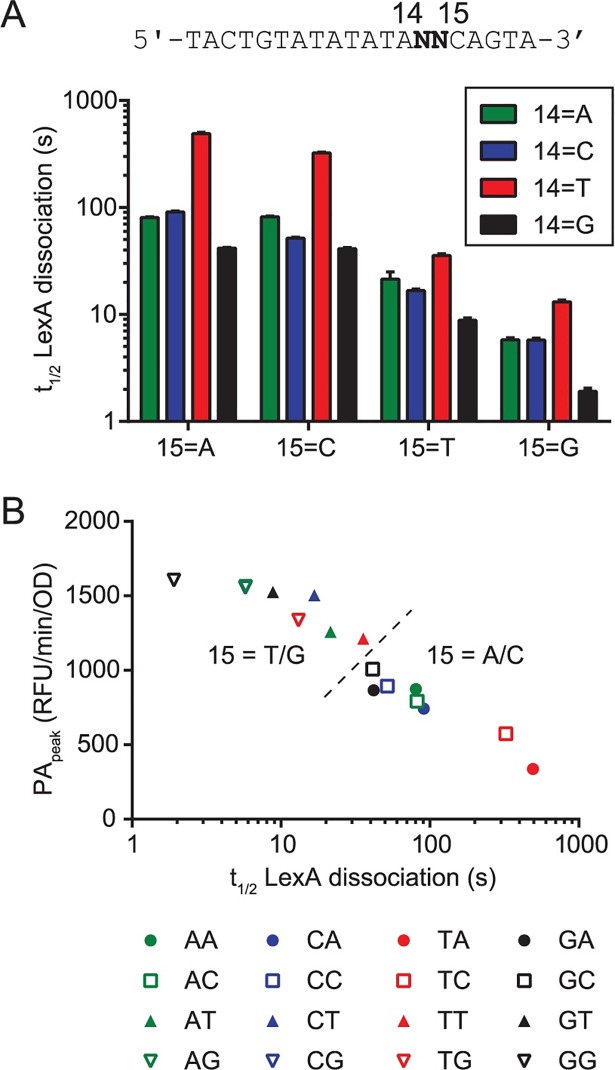
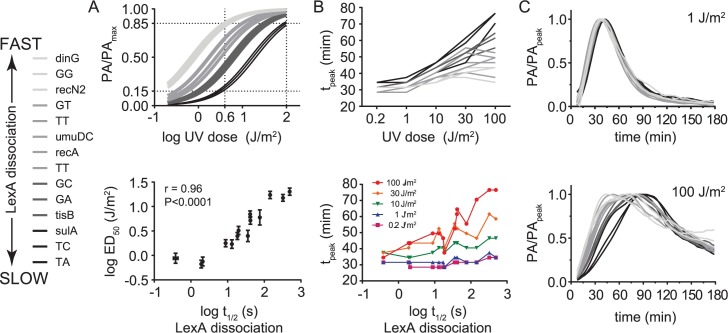
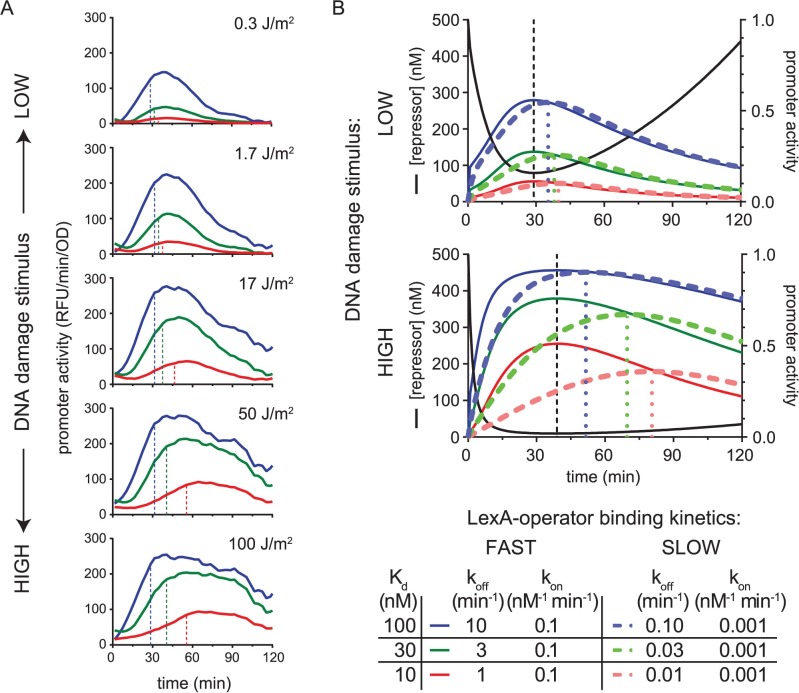
Biochemical pathways are often genetically encoded as simple transcription regulation networks, where one transcription factor regulates the expression of multiple genes in a pathway. The relative timing of each promoter's activation and shut-off within the network can impact physiology. In the DNA damage repair pathway (known as the SOS response) of Escherichia coli, approximately 40 genes are regulated by the LexA repressor. After a DNA damaging event, LexA degradation triggers SOS gene transcription, which is temporally separated into subsets of 'early', 'middle', and 'late' genes. Although this feature plays an important role in regulating the SOS response, both the range of this separation and its underlying mechanism are not experimentally defined. Here we show that, at low doses of DNA damage, the timing of promoter activities is not separated. Instead, timing differences only emerge at higher levels of DNA damage and increase as a function of DNA damage dose. To understand mechanism, we derived a series of synthetic SOS gene promoters which vary in LexA-operator binding kinetics, but are otherwise identical, and then studied their activity over a large dose-range of DNA damage. In distinction to established models based on rapid equilibrium assumptions, the data best fit a kinetic model of repressor occupancy at promoters, where the drop in cellular LexA levels associated with higher doses of DNA damage leads to non-equilibrium binding kinetics of LexA at operators. Operators with slow LexA binding kinetics achieve their minimal occupancy state at later times than operators with fast binding kinetics, resulting in a time separation of peak promoter activity between genes. These data provide insight into this remarkable feature of the SOS pathway by demonstrating how a single transcription factor can be employed to control the relative timing of each gene's transcription as a function of stimulus dose.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Shen-Orr SS, Milo R, Mangan S, Alon U. (2002) Network motifs in the transcriptional regulation network of Escherichia coli. Nat Genet 31(1): 64–68. doi: 10.1038/ng881 - DOI - PubMed
-
- Kalir S, McClure J, Pabbaraju K, Southward C, Ronen M, et al. (2001) Ordering genes in a flagella pathway by analysis of expression kinetics from living bacteria. Science 292(5524): 2080–2083. doi: 10.1126/science.1058758 - DOI - PubMed
-
- Zaslaver A, Mayo AE, Rosenberg R, Bashkin P, Sberro H, et al. (2004) Just-in-time transcription program in metabolic pathways. Nat Genet 36(5): 486–491. doi: 10.1038/ng1348 - DOI - PubMed
-
- Simmons LA, Foti JJ, Cohen SE, Walker GC. (2008) The SOS regulatory network. EcoSal Plus 3(1): doi: 10.1128/ecosalplus.5.4.3 - DOI - PubMed
-
- Radman M. (1999) Enzymes of evolutionary change. Nature 401(6756): 866–7, 869. doi: 10.1038/44738 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- K08 AI127933/AI/NIAID NIH HHS/United States
- T32-AR007442 /NH/NIH HHS/United States
- T32 GM007229/GM/NIGMS NIH HHS/United States
- T32 AR007442/AR/NIAMS NIH HHS/United States
- T32 AI060516/AI/NIAID NIH HHS/United States
- K08-AI127933 /NH/NIH HHS/United States
- DP2 GM105444/GM/NIGMS NIH HHS/United States
- T32-GM7229 /NH/NIH HHS/United States
- DP2-GM105444 /NH/NIH HHS/United States
- R01 GM127593/GM/NIGMS NIH HHS/United States
- R01-GM080279 /NH/NIH HHS/United States
- R01-GM127593 /NH/NIH HHS/United States
- R01 GM080279/GM/NIGMS NIH HHS/United States
- T32-AI060516 /NH/NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
