

Human-Specific NOTCH2NL Genes Expand Cortical Neurogenesis through Delta/Notch Regulation
- PMID: 29856955
- PMCID: PMC6092419
- DOI: 10.1016/j.cell.2018.03.067
Human-Specific NOTCH2NL Genes Expand Cortical Neurogenesis through Delta/Notch Regulation
Abstract
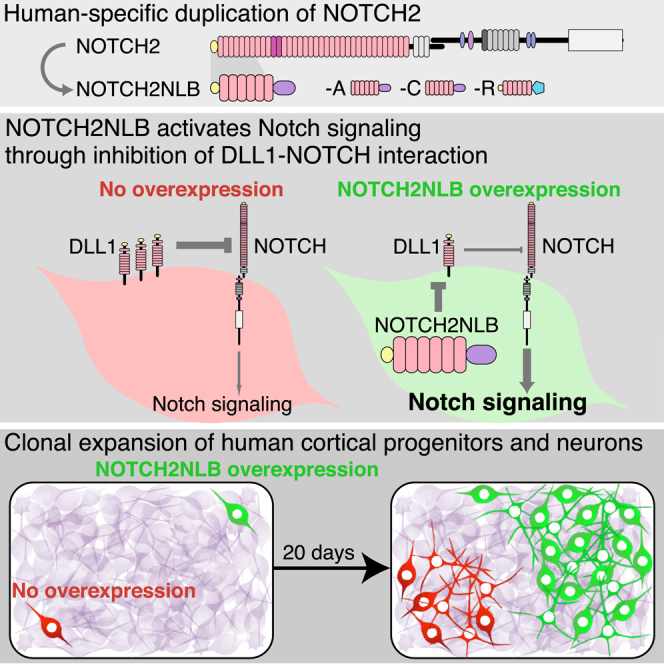
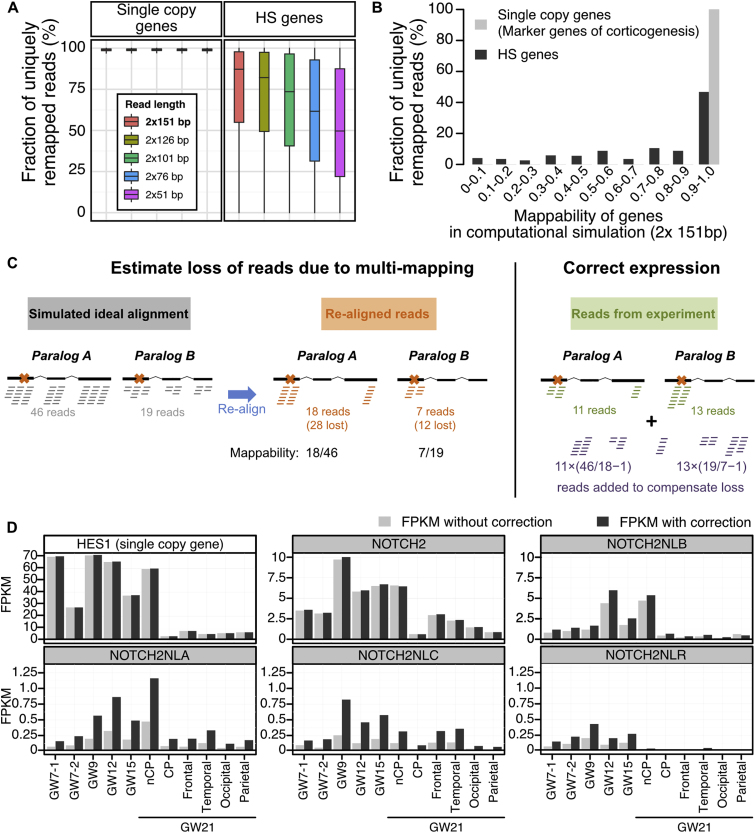
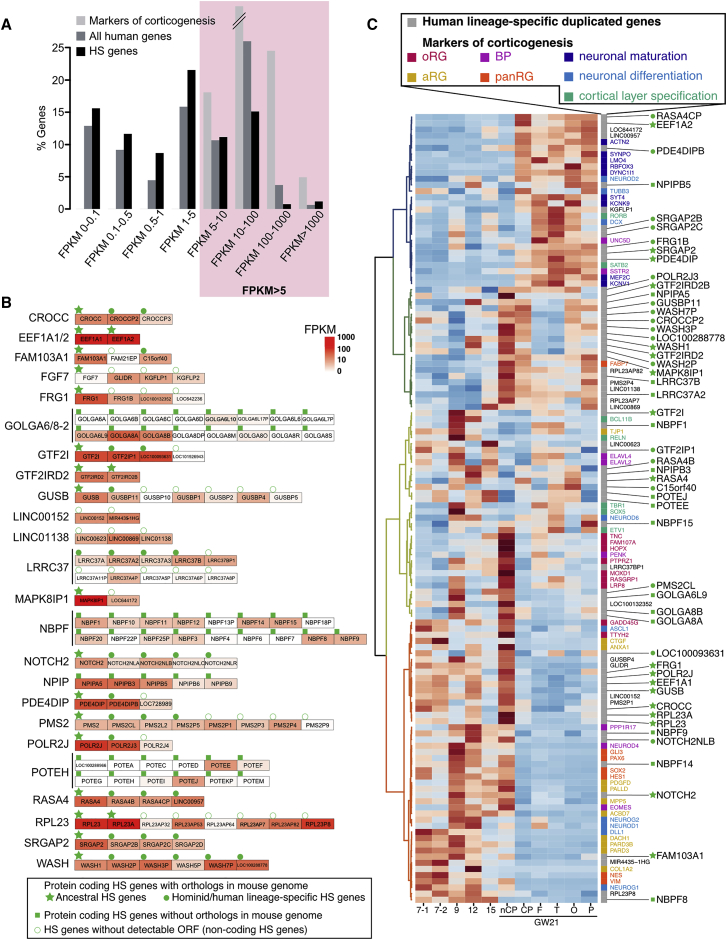
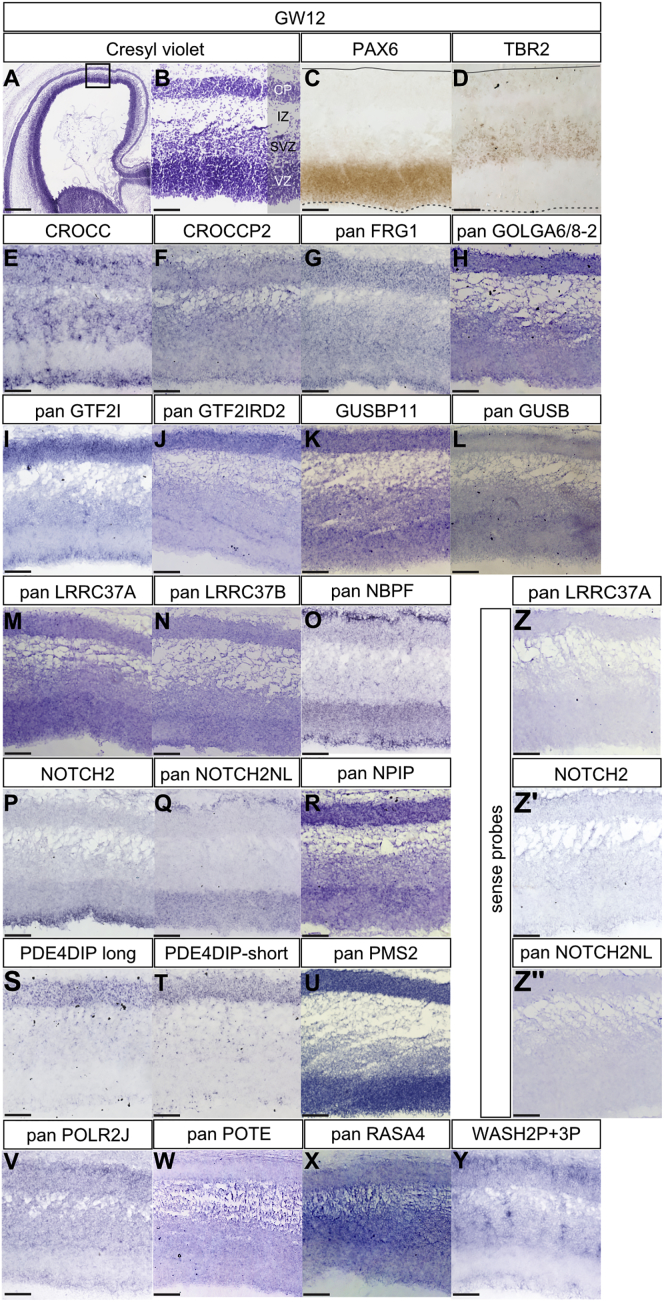
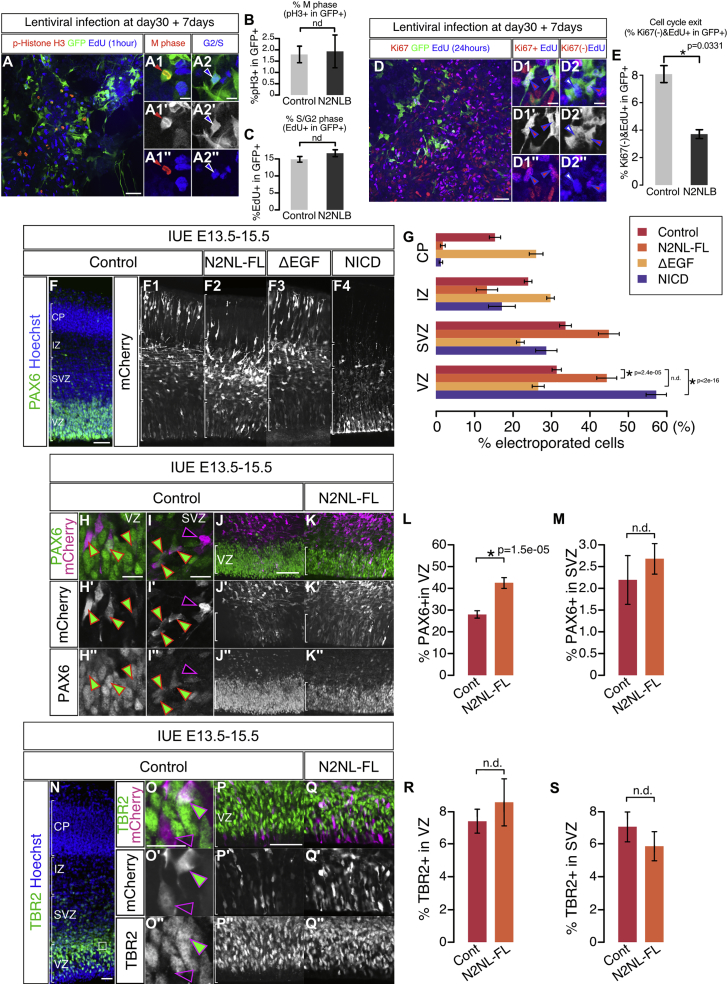
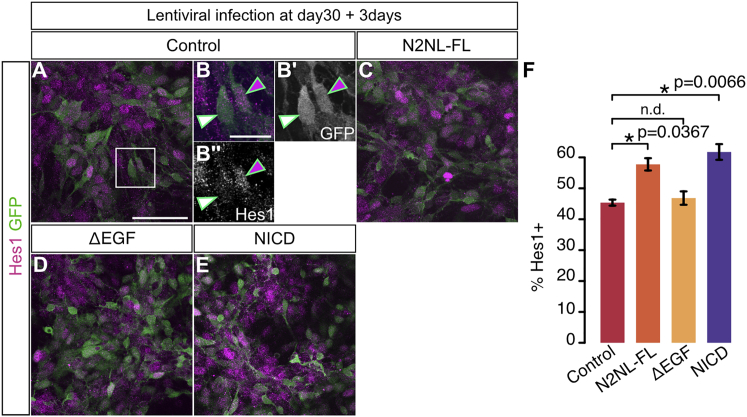
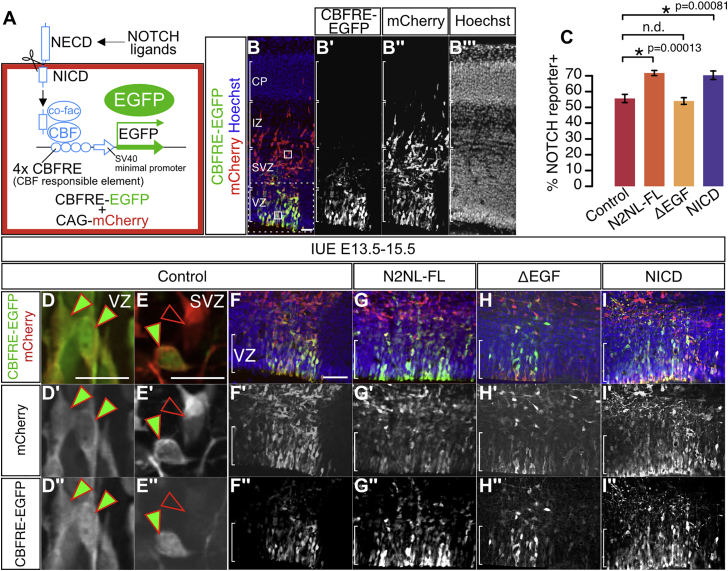
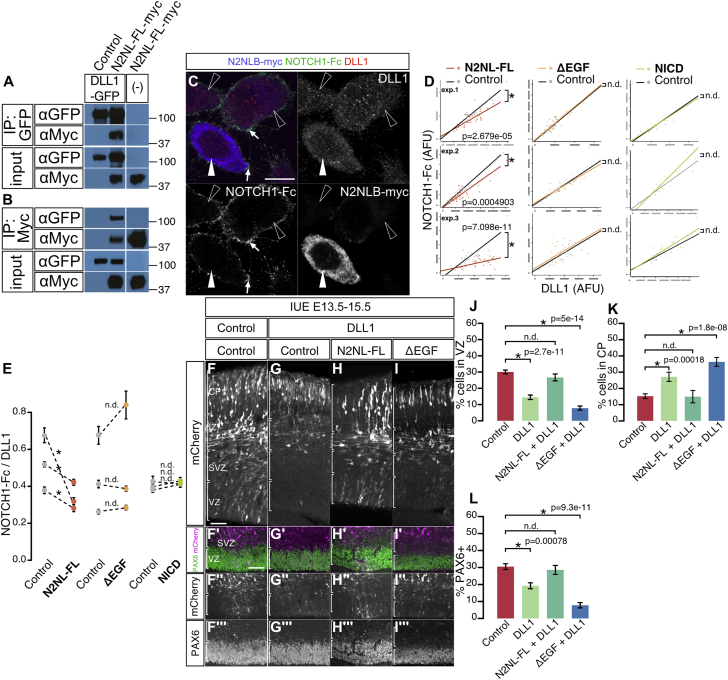
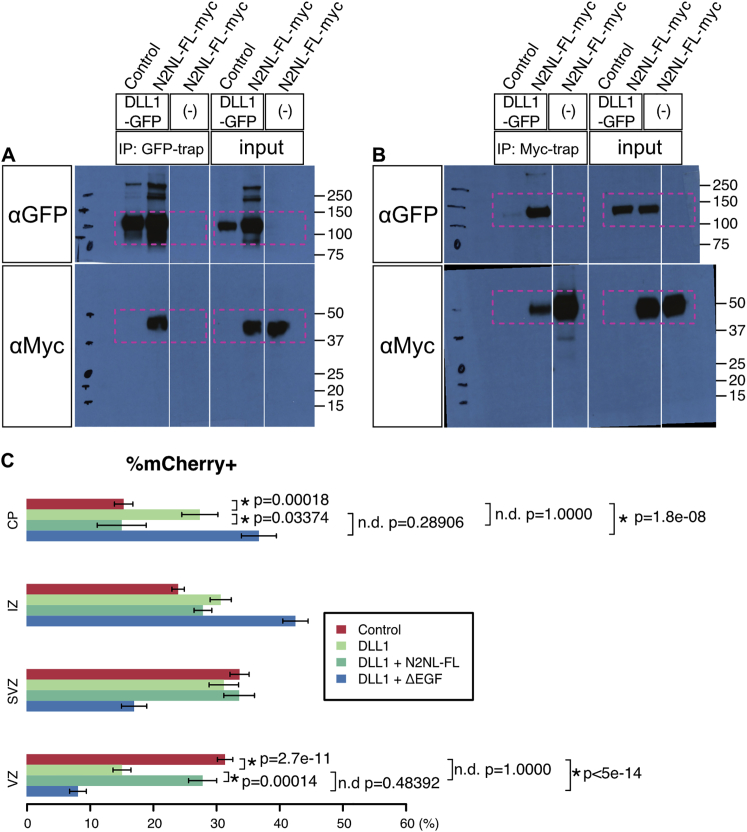
The cerebral cortex underwent rapid expansion and increased complexity during recent hominid evolution. Gene duplications constitute a major evolutionary force, but their impact on human brain development remains unclear. Using tailored RNA sequencing (RNA-seq), we profiled the spatial and temporal expression of hominid-specific duplicated (HS) genes in the human fetal cortex and identified a repertoire of 35 HS genes displaying robust and dynamic patterns during cortical neurogenesis. Among them NOTCH2NL, human-specific paralogs of the NOTCH2 receptor, stood out for their ability to promote cortical progenitor maintenance. NOTCH2NL promote the clonal expansion of human cortical progenitors, ultimately leading to higher neuronal output. At the molecular level, NOTCH2NL function by activating the Notch pathway through inhibition of cis Delta/Notch interactions. Our study uncovers a large repertoire of recently evolved genes active during human corticogenesis and reveals how human-specific NOTCH paralogs may have contributed to the expansion of the human cortex.
Keywords: Notch; brain development; cerebral cortex; human evolution; neurogenesis.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
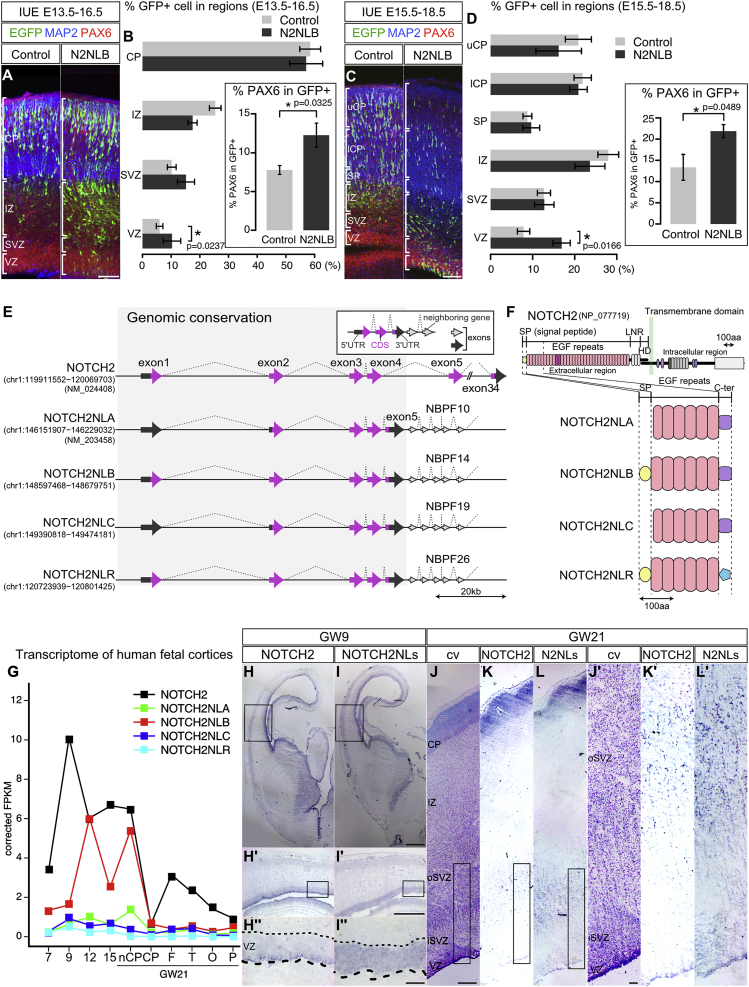
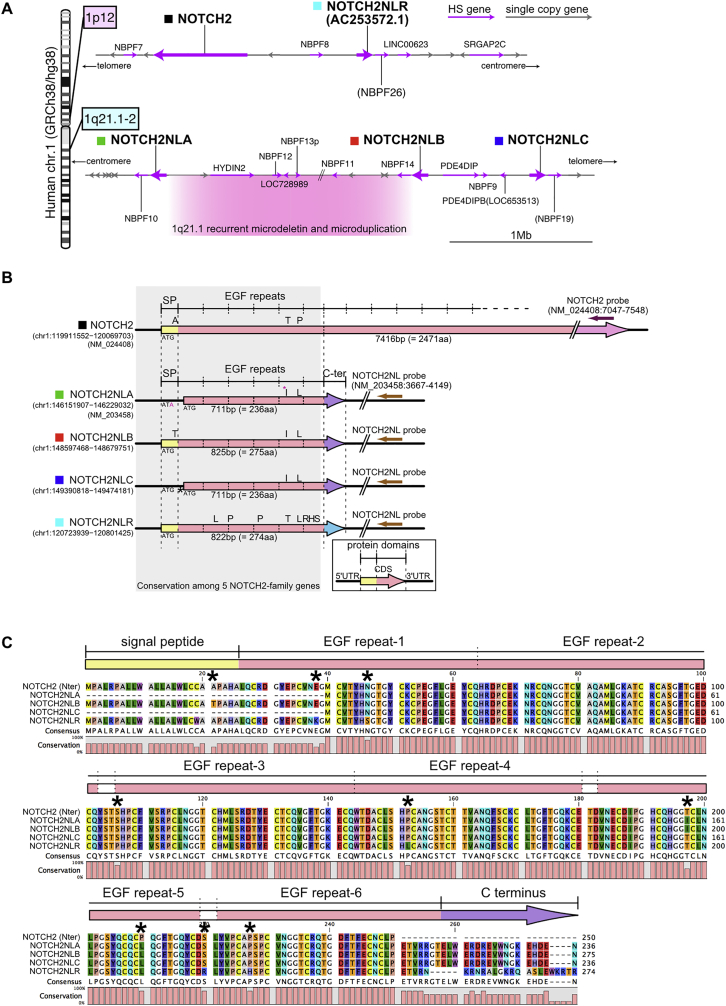
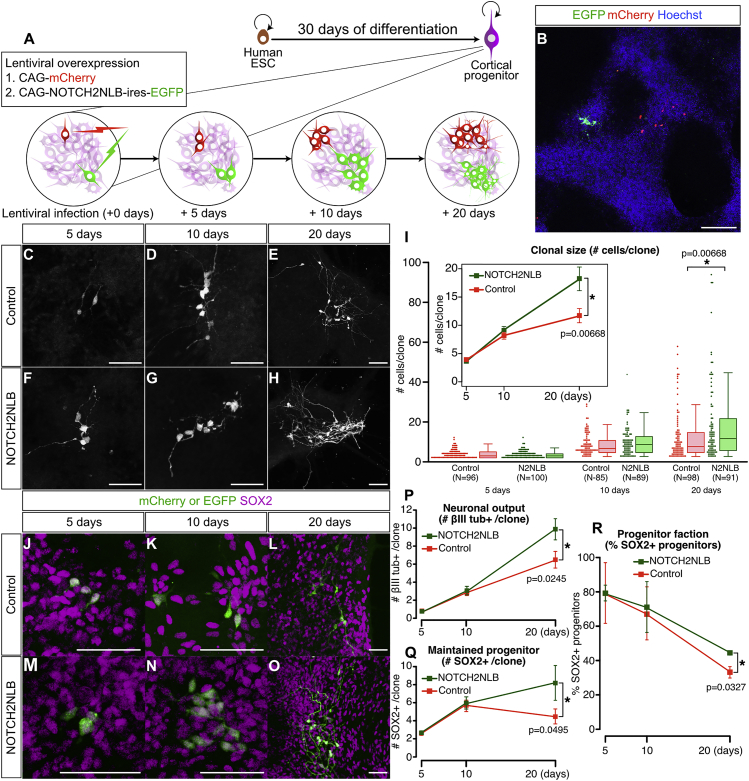
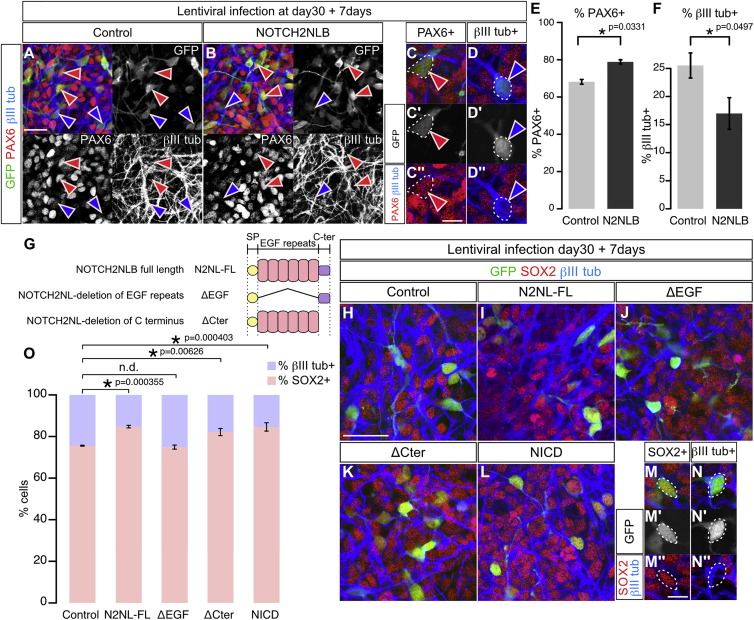
Comment in
-
Making a Notch in the Evolution of the Human Cortex.Dev Cell. 2018 Jun 4;45(5):548-550. doi: 10.1016/j.devcel.2018.05.015. Dev Cell. 2018. PMID: 29870717
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
