

Time-Restricted PiggyBac DNA Transposition by Transposase Protein Delivery Using Lentivirus-Derived Nanoparticles
- PMID: 29858060
- PMCID: PMC5992343
- DOI: 10.1016/j.omtn.2018.02.006
Time-Restricted PiggyBac DNA Transposition by Transposase Protein Delivery Using Lentivirus-Derived Nanoparticles
Abstract
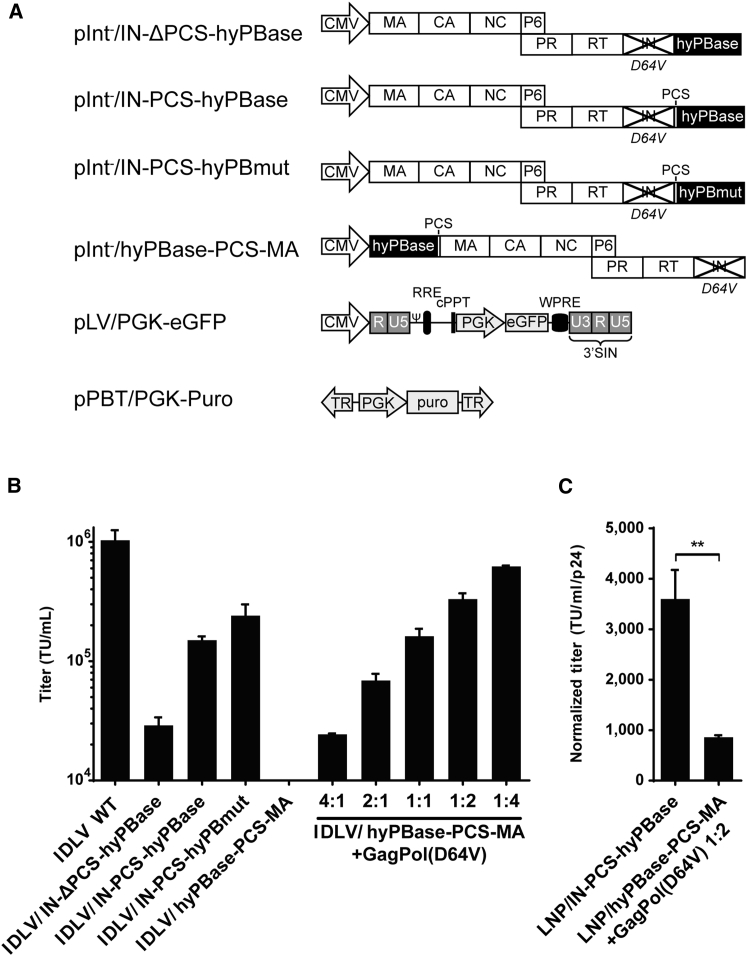
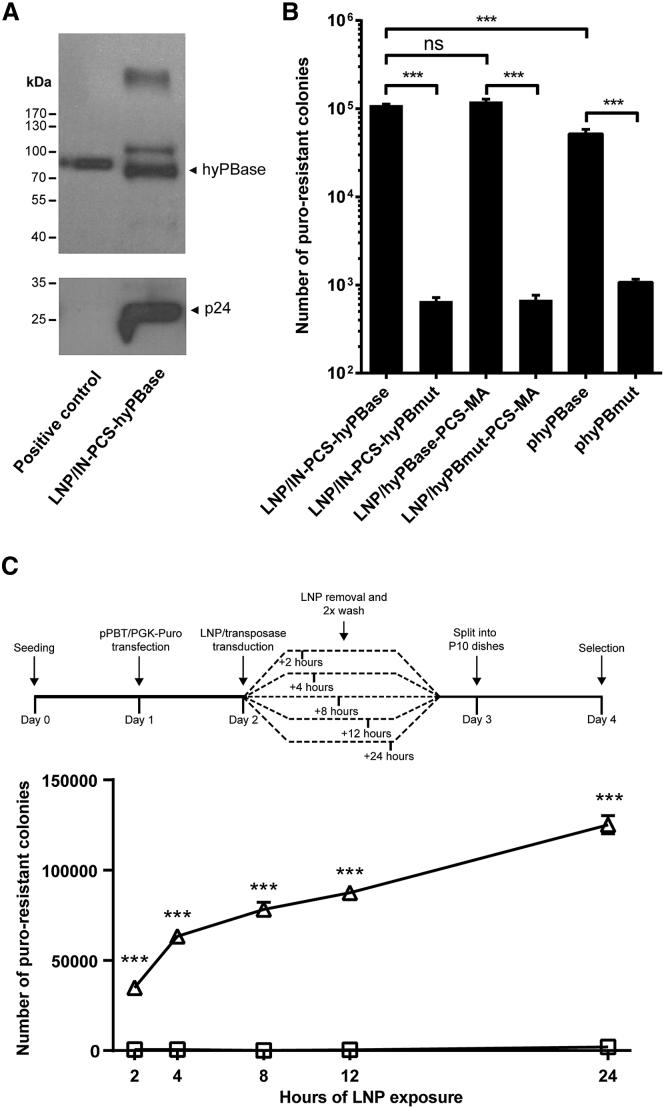
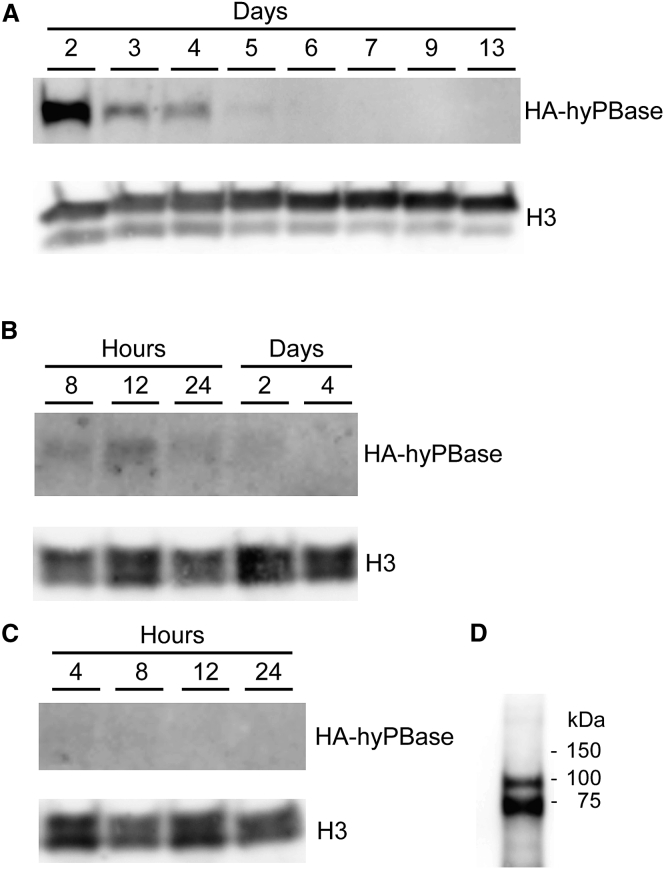
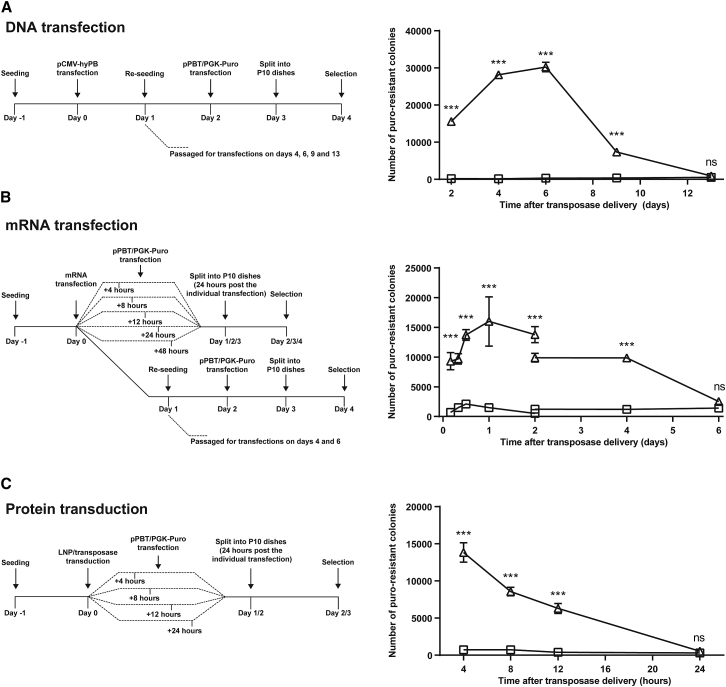
Continuous innovation of revolutionizing genome engineering technologies calls for an intensified focus on new delivery technologies that not only match the inventiveness of genome editors but also enable the combination of potent delivery and time-restricted action of genome-modifying bits and tools. We have previously demonstrated the use of lentivirus-derived nanoparticles (LNPs) as a protein delivery vehicle, incorporating and transferring DNA transposases, designer nucleases, or RNA-guided endonucleases fused to the N terminus of the Gag/GagPol polypeptide. Here, we establish LNP-directed transfer of the piggyBac DNA transposase protein by fusing the transposase to the integrase protein in the C-terminal end of GagPol. We show protein incorporation and proteolytic release of the DNA transposase within matured LNPs, resulting in high levels of DNA transposition activity in LNP-treated cells. Importantly, as opposed to conventional delivery methods based on transfection of plasmid DNA or in-vitro-transcribed mRNA, protein delivery by LNPs effectively results in time-restricted action of the protein (<24 hr) without compromising overall potency. Our findings refine LNP-directed piggyBac transposase delivery, at present the only available direct delivery strategy for this particular protein, and demonstrate a novel strategy for restricting and fine-tuning the exposure of the genome to DNA-modifying enzymes.
Keywords: DNA transposon; LNP; lentivirus; piggyBac; protein delivery; protein transduction.
Copyright © 2018 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
