

Avidity of α-fucose on human milk oligosaccharides and blood group-unrelated oligo/polyfucoses is essential for potent norovirus-binding targets
- PMID: 29858242
- PMCID: PMC6066315
- DOI: 10.1074/jbc.RA117.001369
Avidity of α-fucose on human milk oligosaccharides and blood group-unrelated oligo/polyfucoses is essential for potent norovirus-binding targets
Abstract
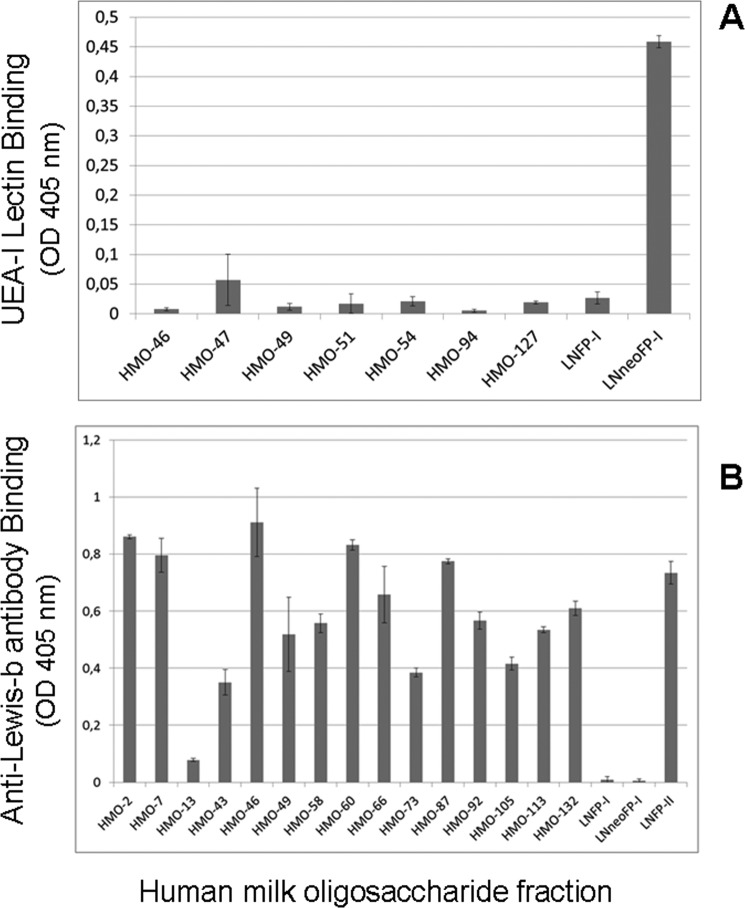
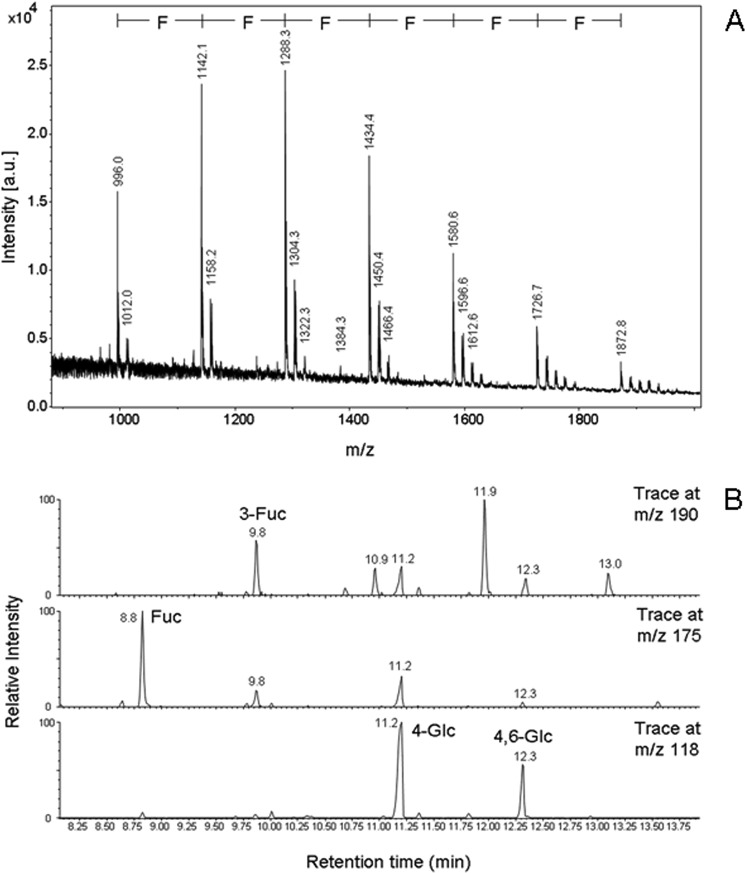
There is agreement with respect to norovirus infection routes in humans regarding binding of the pathogen to gastrointestinal epithelia via recognition of blood group-active mucin-typeO-glycans as the initiating and essential event. Among food additives playing a potential role in applications to protect newborns, human milk oligosaccharides (HMOs) as competitors are of major importance. By focusing on fractions of high-molecular mass HMOs with high fucose contents, we attempted to identify the structural elements required for norovirus GII.4 (Sydney 2012, JX459908) capsid binding in neoglycolipid-based arrays. We provide evidence that HMO fractions with the strongest binding capacities contained hepta- to decasaccharides expressing branches with terminal blood group H1 or Lewis-b antigen. H2 antigen, as recognized by UEA-I lectin, is apparently not expressed in high-mass HMOs. Beyond affinity, sterical and valency effects contribute more to virus-like particle binding, as revealed for oligovalent fucose conjugates of α-cyclodextrin and oligofucoses from fucoidan. Accordingly, high-mass HMOs with oligovalent fucose can exhibit stronger binding capacities compared with monovalent fucose HMOs. The above features were revealed for the most clinically relevant and prevalent GII.4 strain and are distinct from other strains, like GII.10 (Vietnam 026, AF504671), which showed a preference for blood group Lewis-a positive glycans.
Keywords: anti-viral protection; carbohydrate-binding protein; food additive; glycobiology; human blood group antigen; innate immunity; milk oligosaccharide; norovirus; oligosaccharide; virus.
© 2018 by The American Society for Biochemistry and Molecular Biology, Inc.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures







References
-
- van Beek J., Ambert-Balay K., Botteldoorn N., Eden J. S., Fonager J., Hewitt J., Iritani N., Kroneman A., Vennema H., Vinjé J., White P. A., Koopmans M., and NoroNet. (2013) Indications for worldwide increased norovirus activity associated with emergence of a new variant of genotype II.4, late 2012. Euro Surveill. 18, 8–9 - PubMed
-
- Fankhauser R. L., Monroe S. S., Noel J. S., Humphrey C. D., Bresee J. S., Parashar U. D., Ando T., and Glass R. I. (2002) Epidemiologic and molecular trends of “Norwalk-like Viruses” associated with outbreaks of gastroenteritis in the United States. J. Infect. Dis. 186, 1–7 10.1086/341085 - DOI - PubMed
-
- Marionneau S., Ruvoën N., Le Moullac-Vaidye B., Clement M., Cailleau-Thomas A., Ruiz-Palacois G., Huang P., Jiang X., and Le Pendu J. (2002) Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 122, 1967–1977 10.1053/gast.2002.33661 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
