

Cellular conditions of weakly chelated magnesium ions strongly promote RNA stability and catalysis
- PMID: 29858572
- PMCID: PMC5984629
- DOI: 10.1038/s41467-018-04415-1
Cellular conditions of weakly chelated magnesium ions strongly promote RNA stability and catalysis
Abstract
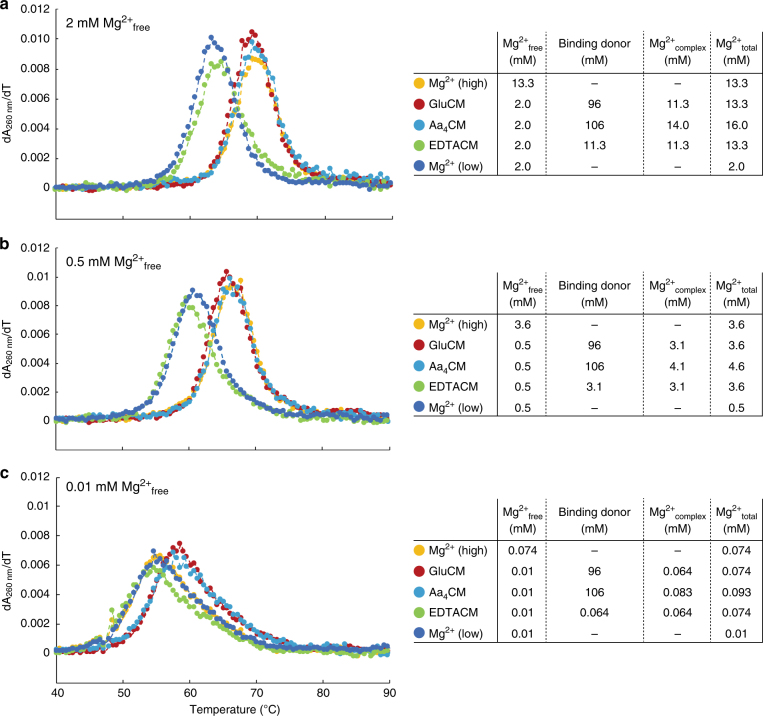
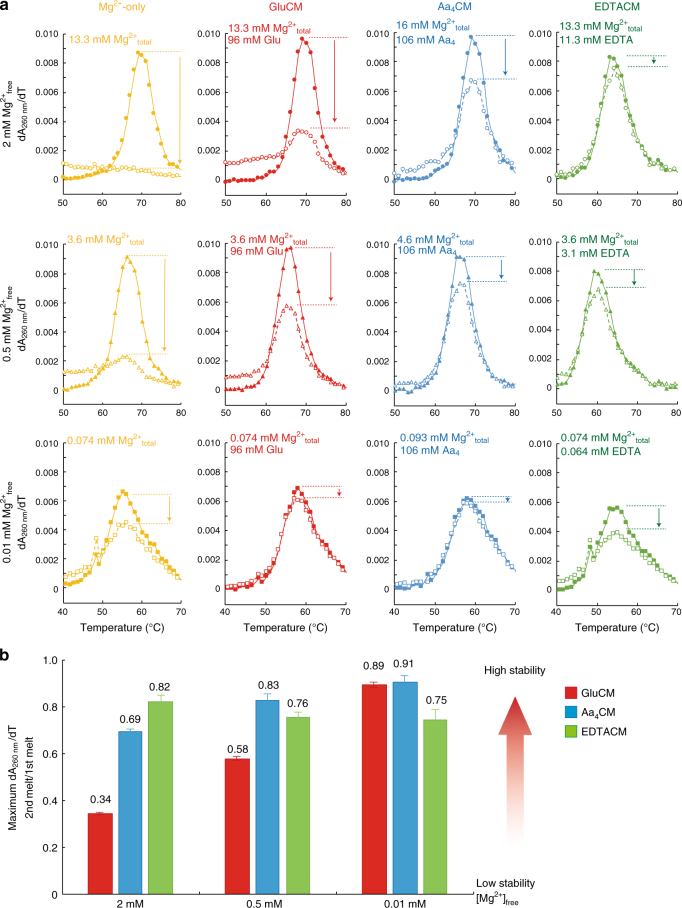
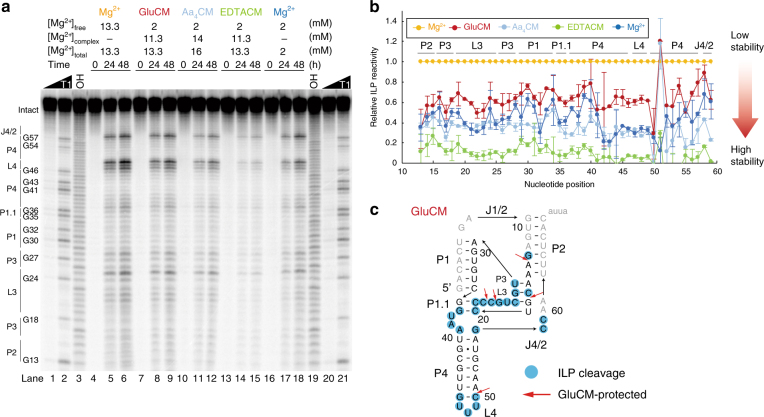
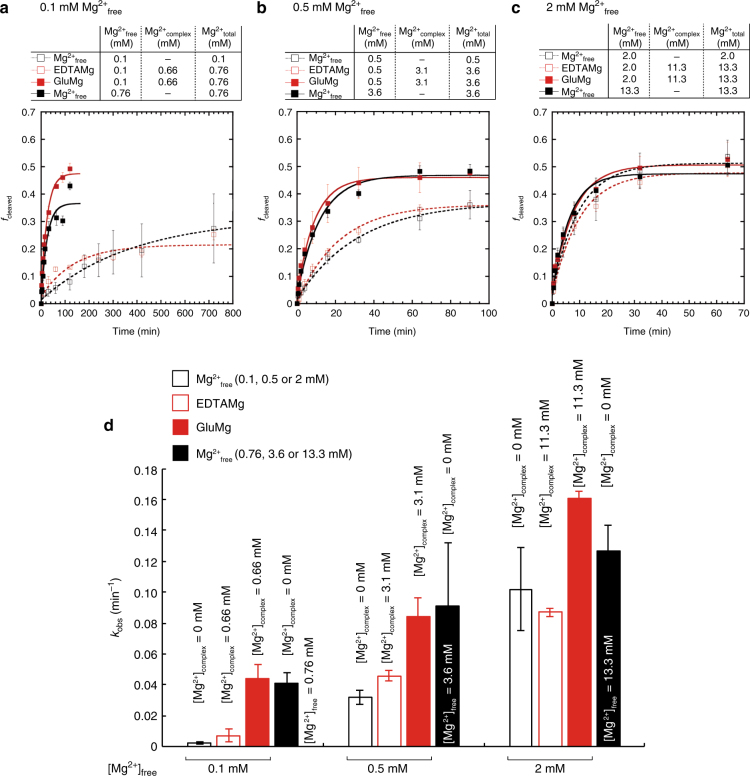
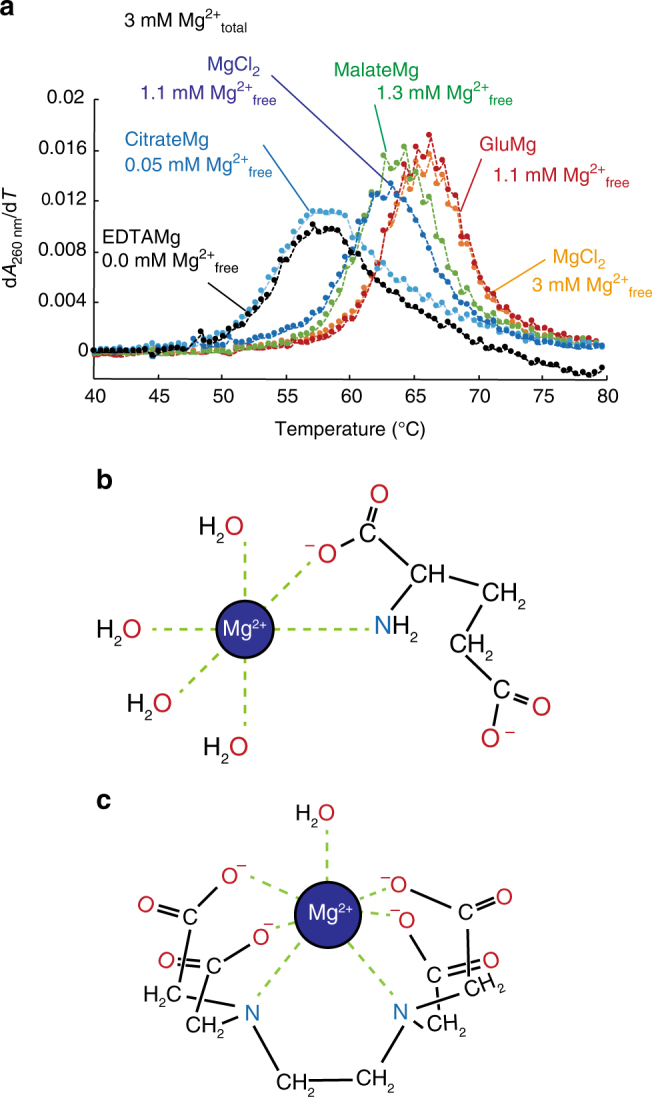
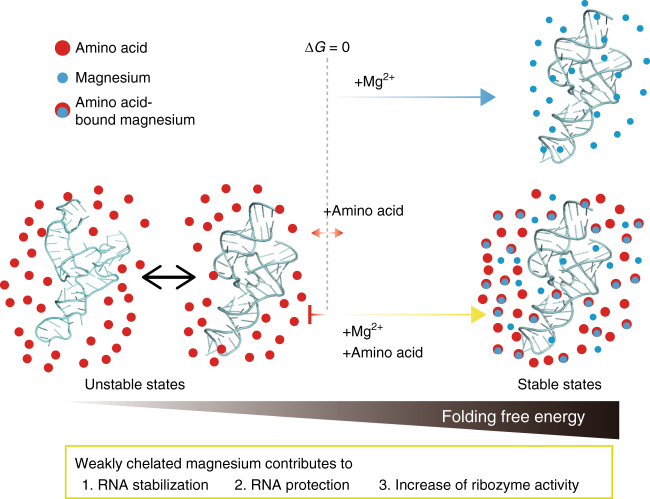
Most RNA folding studies have been performed under non-physiological conditions of high concentrations (≥10 mM) of Mg2+free, while actual cellular concentrations of Mg2+free are only ~1 mM in a background of greater than 50 mM Mg2+total. To uncover cellular behavior of RNA, we devised cytoplasm mimic systems that include biological concentrations of amino acids, which weakly chelate Mg2+. Amino acid-chelated Mg2+ (aaCM) of ~15 mM dramatically increases RNA folding and prevents RNA degradation. Furthermore, aaCM enhance self-cleavage of several different ribozymes, up to 100,000-fold at Mg2+free of just 0.5 mM, indirectly through RNA compaction. Other metabolites that weakly chelate magnesium offer similar beneficial effects, which implies chelated magnesium may enhance RNA function in the cell in the same way. Overall, these results indicate that the states of Mg2+ should not be limited to free and bound only, as weakly bound Mg2+ strongly promotes RNA function under cellular conditions.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
