

Control of Feeding Behavior by Cerebral Ventricular Volume Transmission of Melanin-Concentrating Hormone
- PMID: 29861386
- PMCID: PMC6400641
- DOI: 10.1016/j.cmet.2018.05.001
Control of Feeding Behavior by Cerebral Ventricular Volume Transmission of Melanin-Concentrating Hormone
Abstract
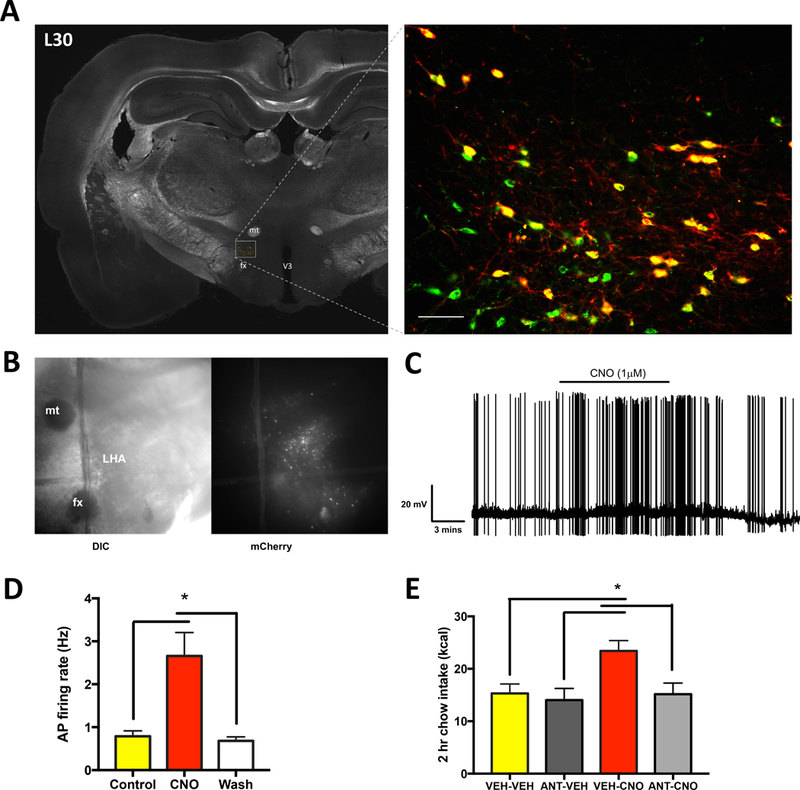
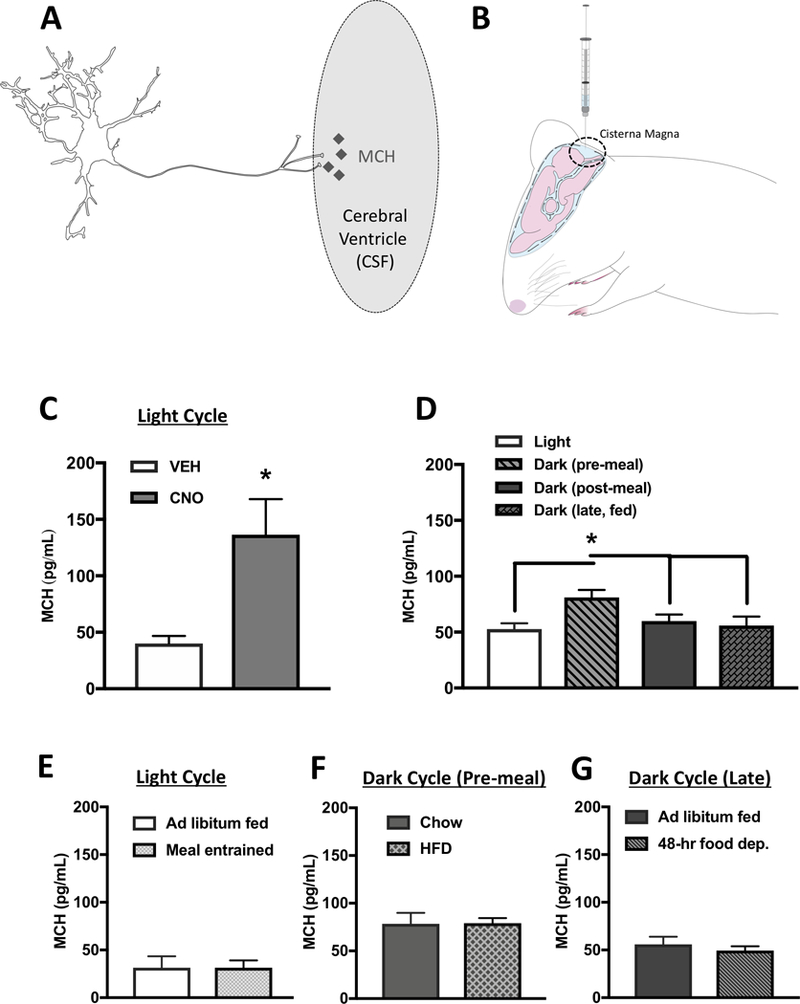
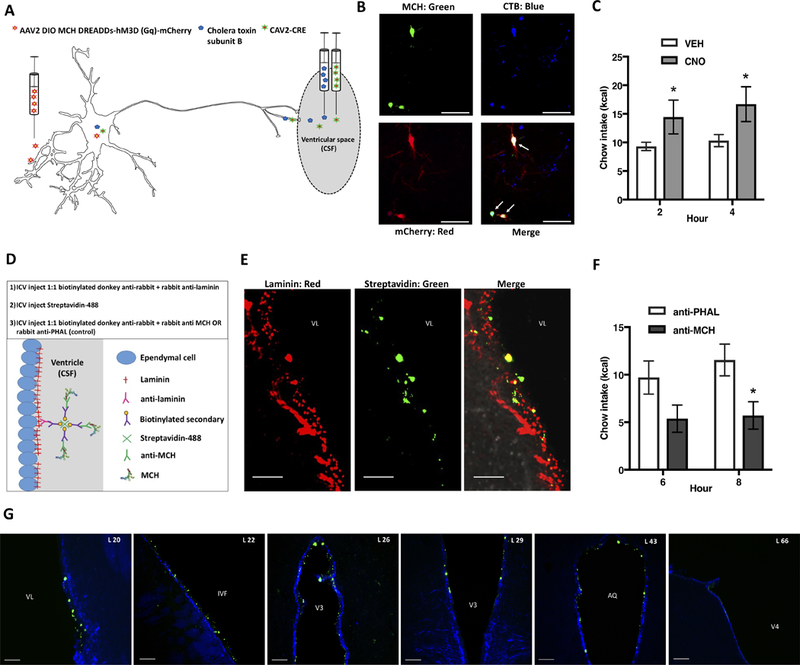
Classical mechanisms through which brain-derived molecules influence behavior include neuronal synaptic communication and neuroendocrine signaling. Here we provide evidence for an alternative neural communication mechanism that is relevant for food intake control involving cerebroventricular volume transmission of the neuropeptide melanin-concentrating hormone (MCH). Results reveal that the cerebral ventricles receive input from approximately one-third of MCH-producing neurons. Moreover, MCH cerebrospinal fluid (CSF) levels increase prior to nocturnal feeding and following chemogenetic activation of MCH-producing neurons. Utilizing a dual viral vector approach, additional results reveal that selective activation of putative CSF-projecting MCH neurons increases food intake. In contrast, food intake was reduced following immunosequestration of MCH endogenously present in CSF, indicating that neuropeptide transmission through the cerebral ventricles is a physiologically relevant signaling pathway for energy balance control. Collectively these results suggest that neural-CSF volume transmission signaling may be a common neurobiological mechanism for the control of fundamental behaviors.
Keywords: CSF; MCH; appetite; cerebrospinal fluid; circadian; feeding; neuroendocrine; obesity; orexin; volume transmission.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
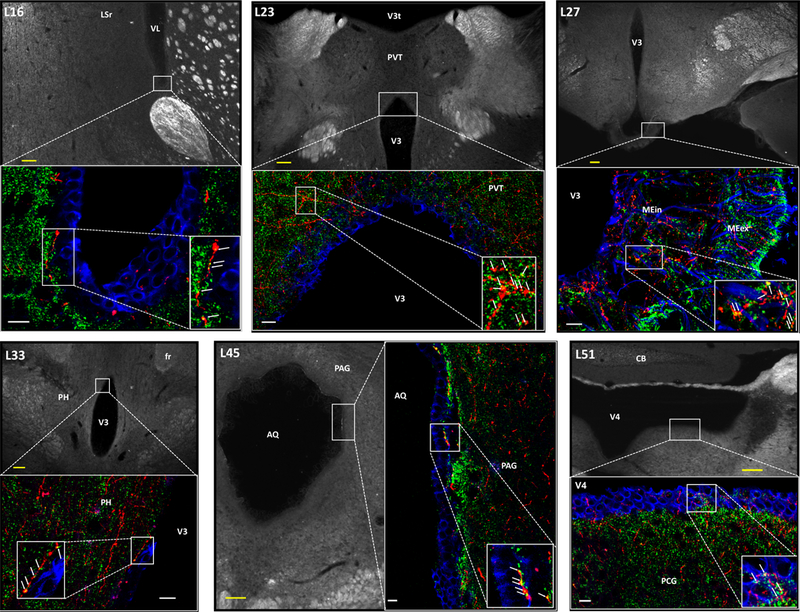
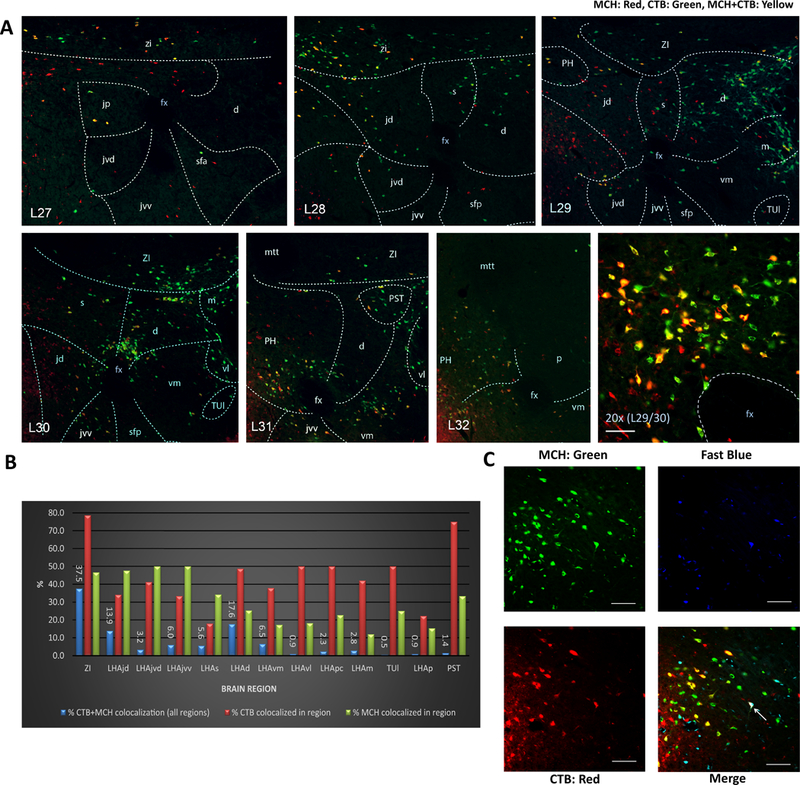
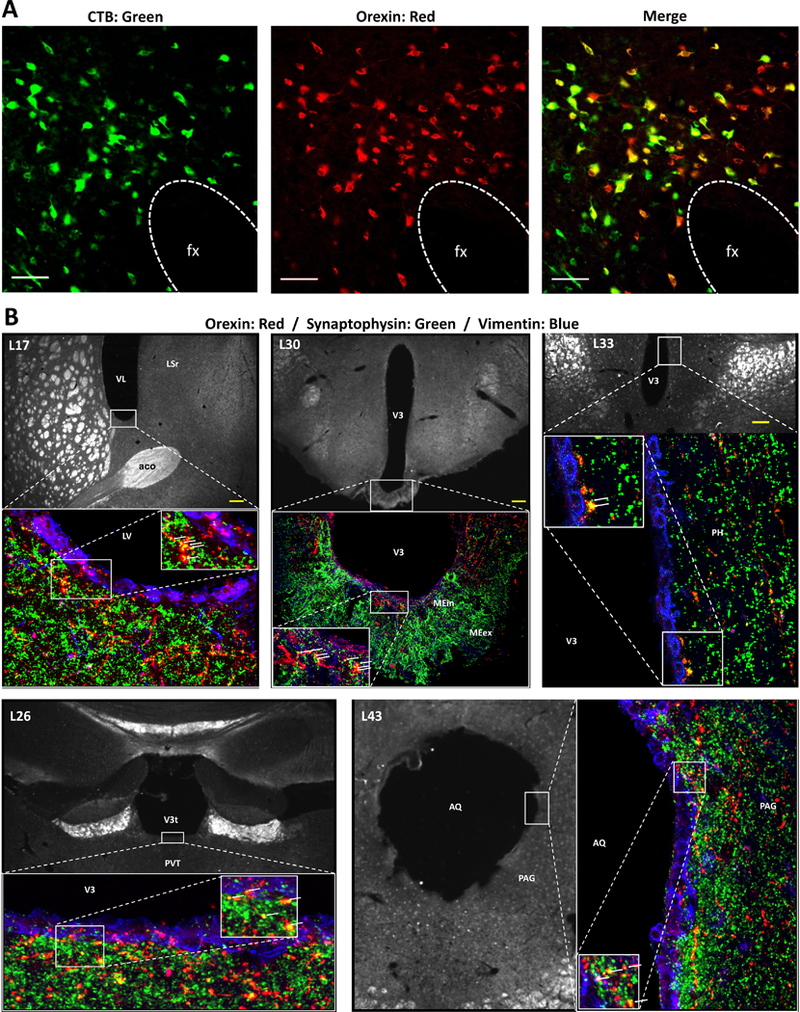
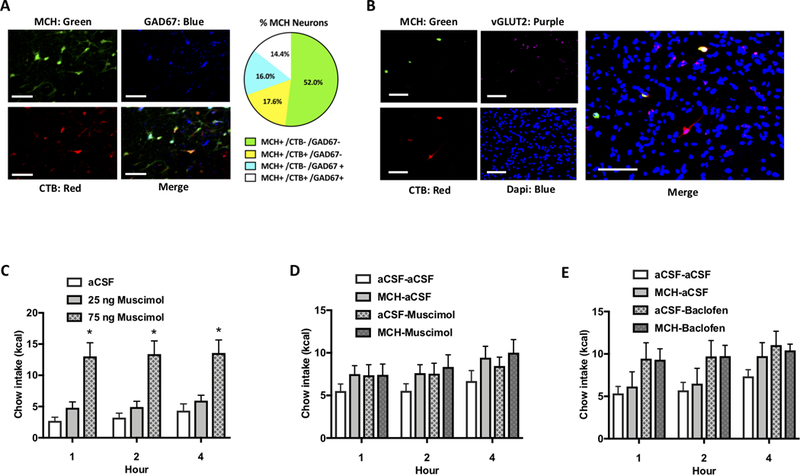
Comment in
-
A novel pathway that controls feeding behaviour.Nat Rev Endocrinol. 2018 Aug;14(8):442. doi: 10.1038/s41574-018-0047-8. Nat Rev Endocrinol. 2018. PMID: 29915219 No abstract available.
-
Melanin-Concentrating Hormone-Dependent Control of Feeding: When Volume Matters.Cell Metab. 2018 Jul 3;28(1):7-8. doi: 10.1016/j.cmet.2018.06.018. Cell Metab. 2018. PMID: 29972799
References
-
- Agnati LF, Fuxe K, Zoli M, Ozini I, Toffano G, and Ferraguti F (1986). A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive terminals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta physiologica Scandinavica 128, 201–207. - PubMed
-
- Bittencourt JC, Presse F, Arias C, Peto C, Vaughan J, Nahon JL, Vale W, and Sawchenko PE (1992). The melanin-concentrating hormone system of the rat brain: an immuno- and hybridization histochemical characterization. J Comp Neurol 319, 218–245. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
