

Sphingolipid biosynthesis in man and microbes
- PMID: 29863195
- PMCID: PMC6148460
- DOI: 10.1039/c8np00019k
Sphingolipid biosynthesis in man and microbes
Abstract
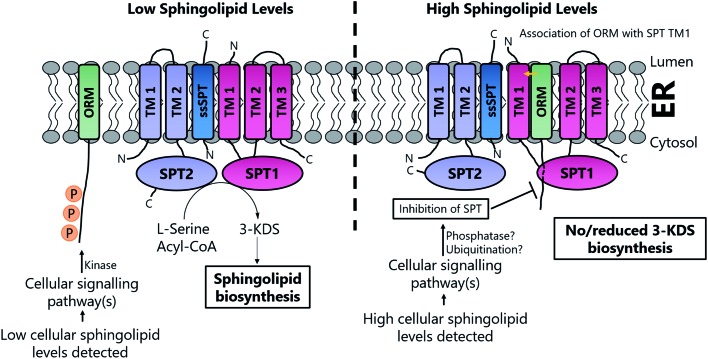
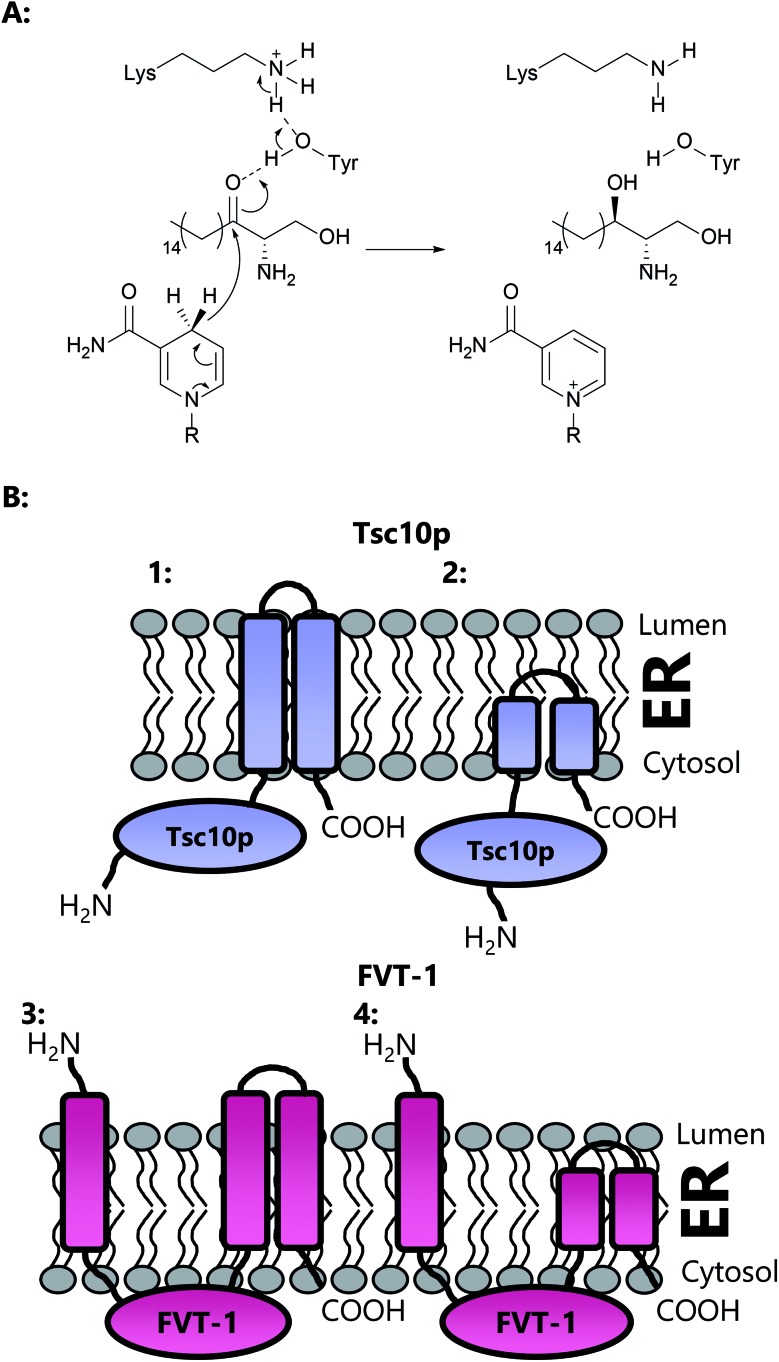
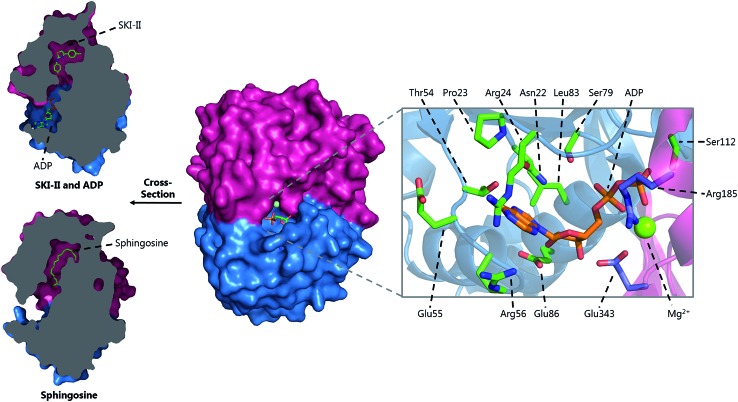
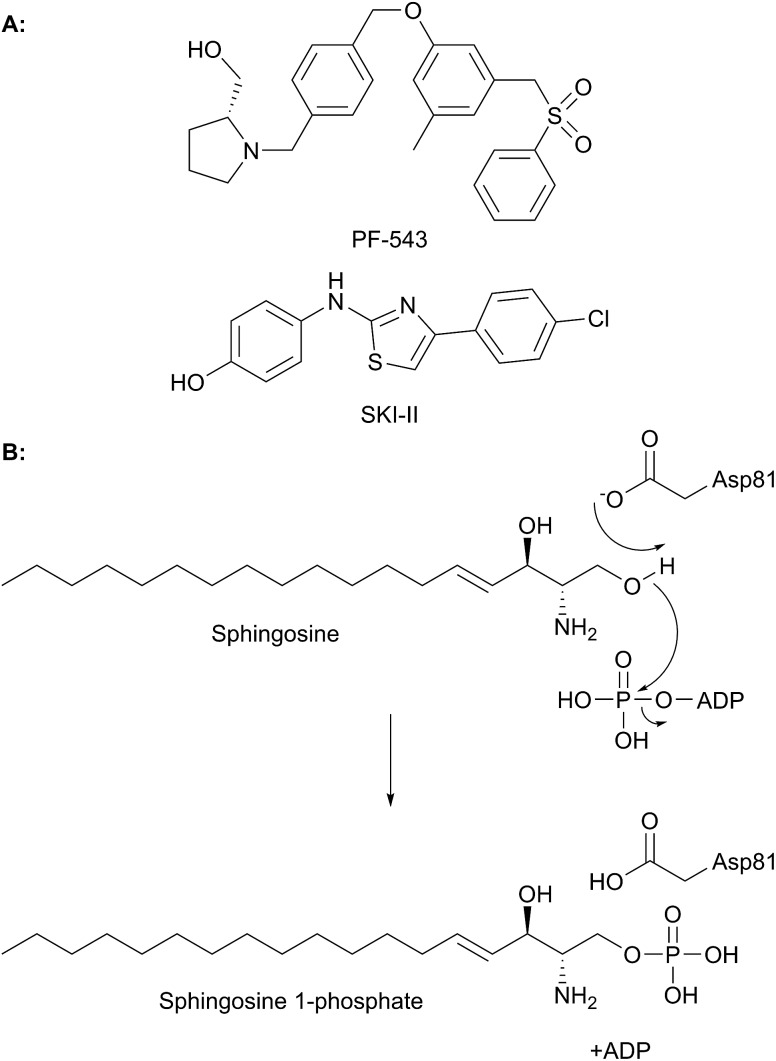
A new review covering up to 2018 Sphingolipids are essential molecules that, despite their long history, are still stimulating interest today. The reasons for this are that, as well as playing structural roles within cell membranes, they have also been shown to perform a myriad of cell signalling functions vital to the correct function of eukaryotic and prokaryotic organisms. Indeed, sphingolipid disregulation that alters the tightly-controlled balance of these key lipids has been closely linked to a number of diseases such as diabetes, asthma and various neuropathologies. Sphingolipid biogenesis, metabolism and regulation is mediated by a large number of enzymes, proteins and second messengers. There appears to be a core pathway common to all sphingolipid-producing organisms but recent studies have begun to dissect out important, species-specific differences. Many of these have only recently been discovered and in most cases the molecular and biochemical details are only beginning to emerge. Where there is a direct link from classic biochemistry to clinical symptoms, a number a drug companies have undertaken a medicinal chemistry campaign to try to deliver a therapeutic intervention to alleviate a number of diseases. Where appropriate, we highlight targets where natural products have been exploited as useful tools. Taking all these aspects into account this review covers the structural, mechanistic and regulatory features of sphingolipid biosynthetic and metabolic enzymes.
Figures
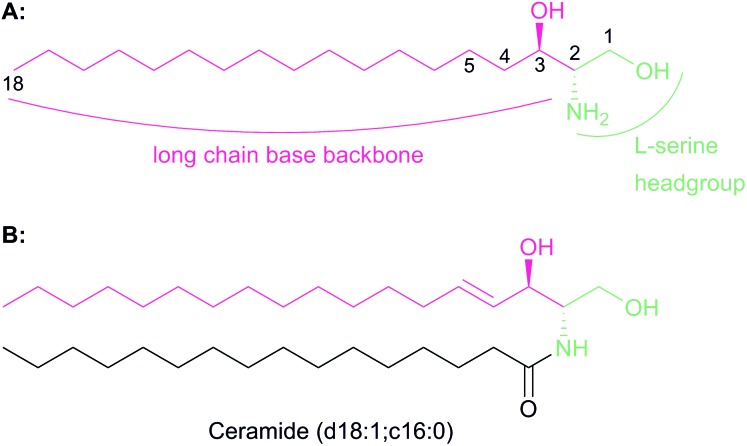
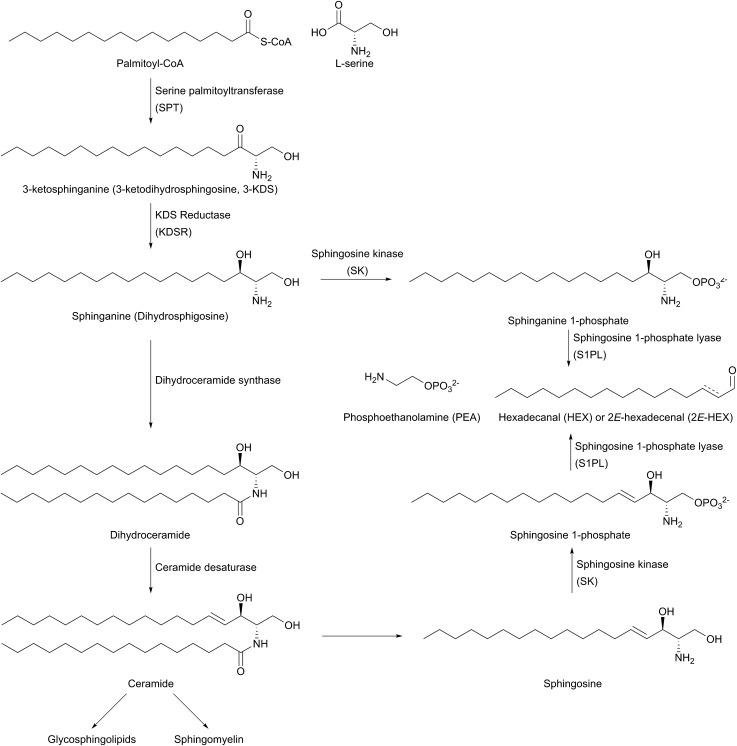
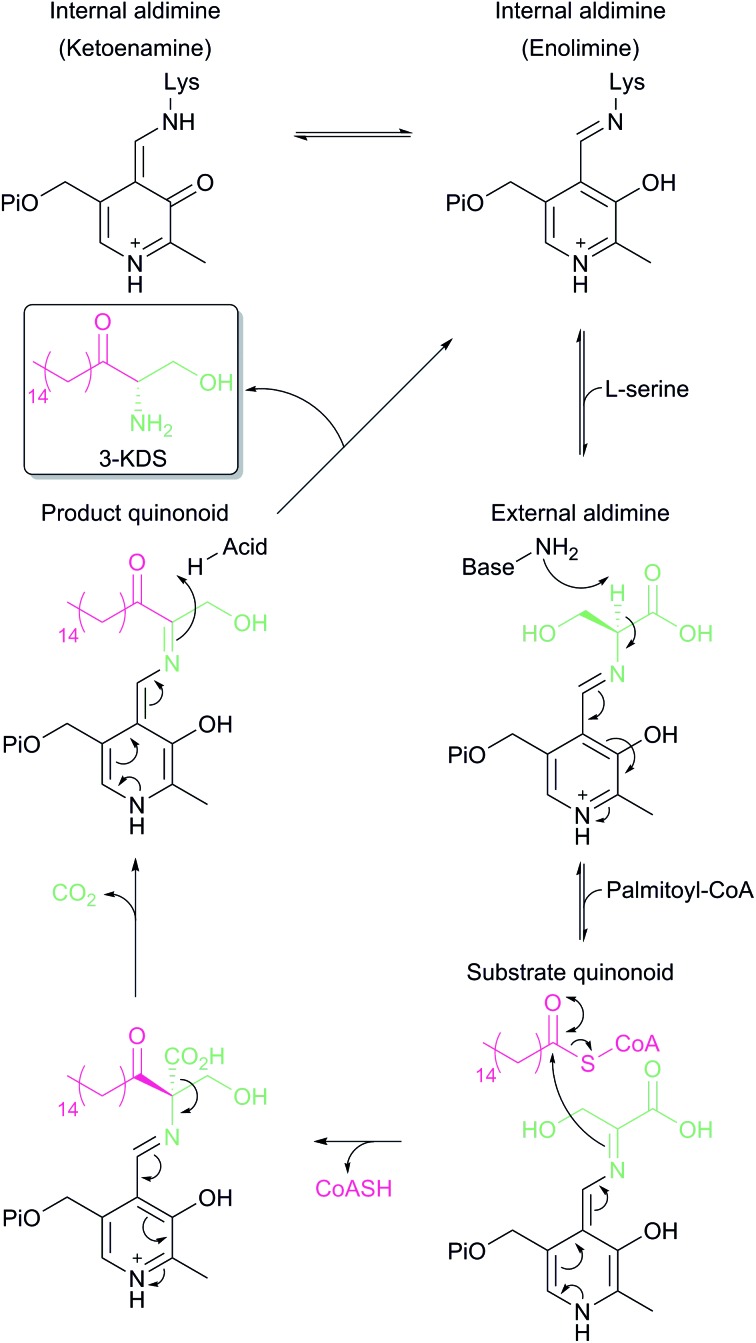
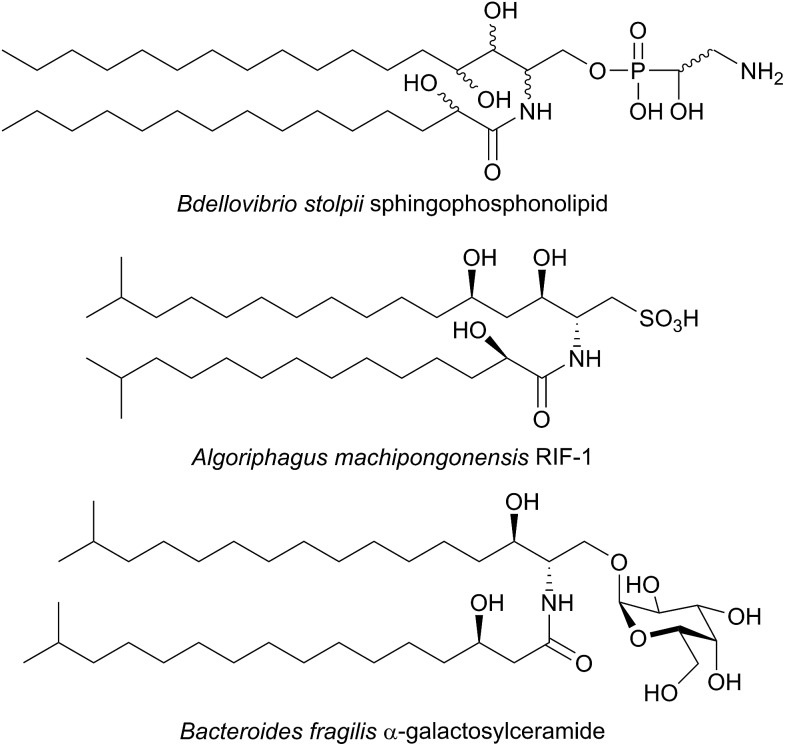
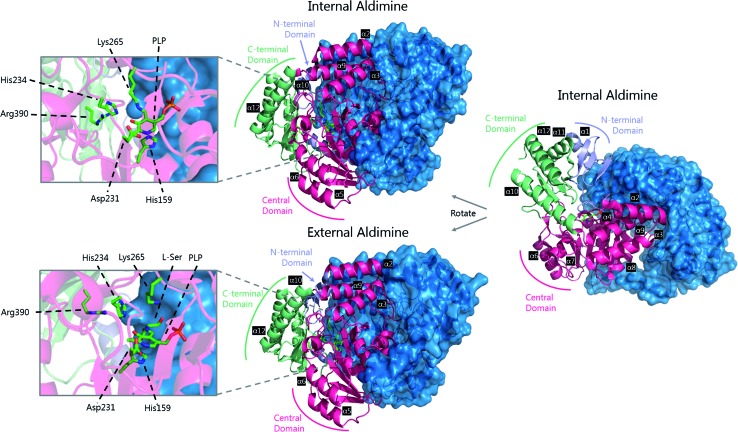
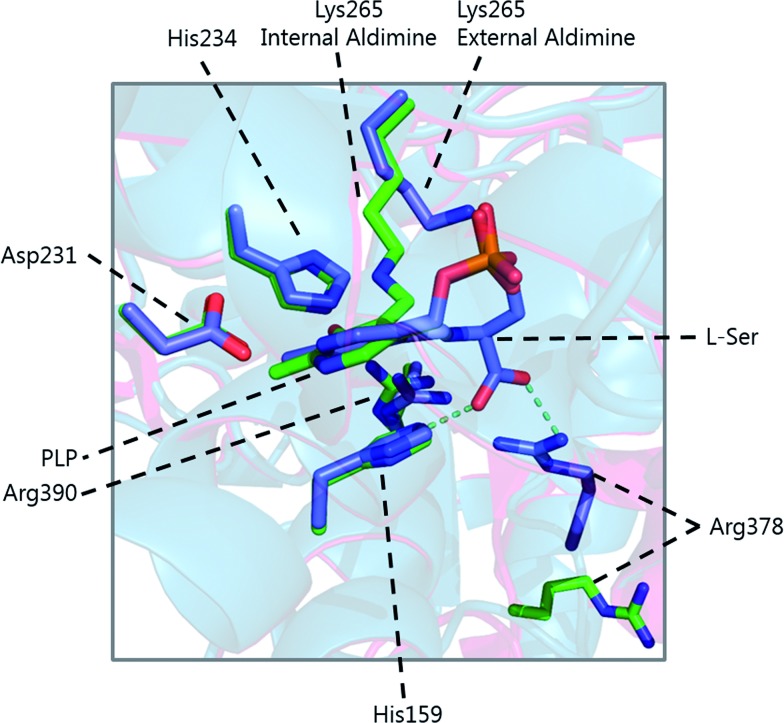

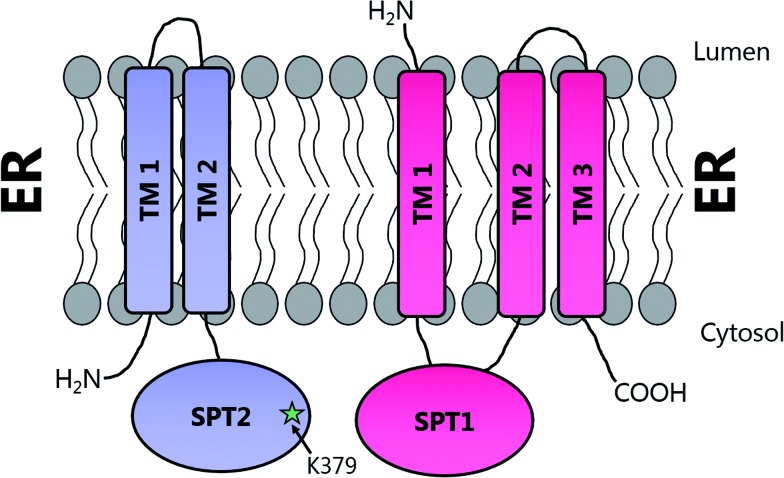
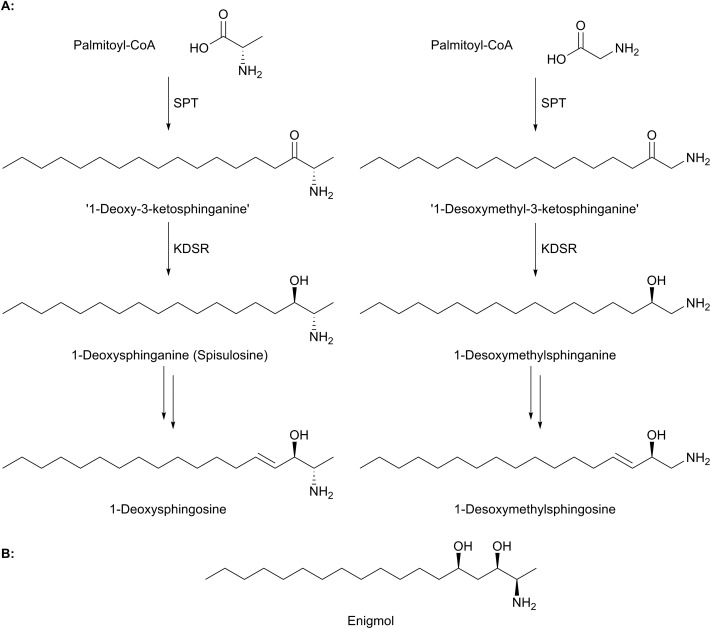
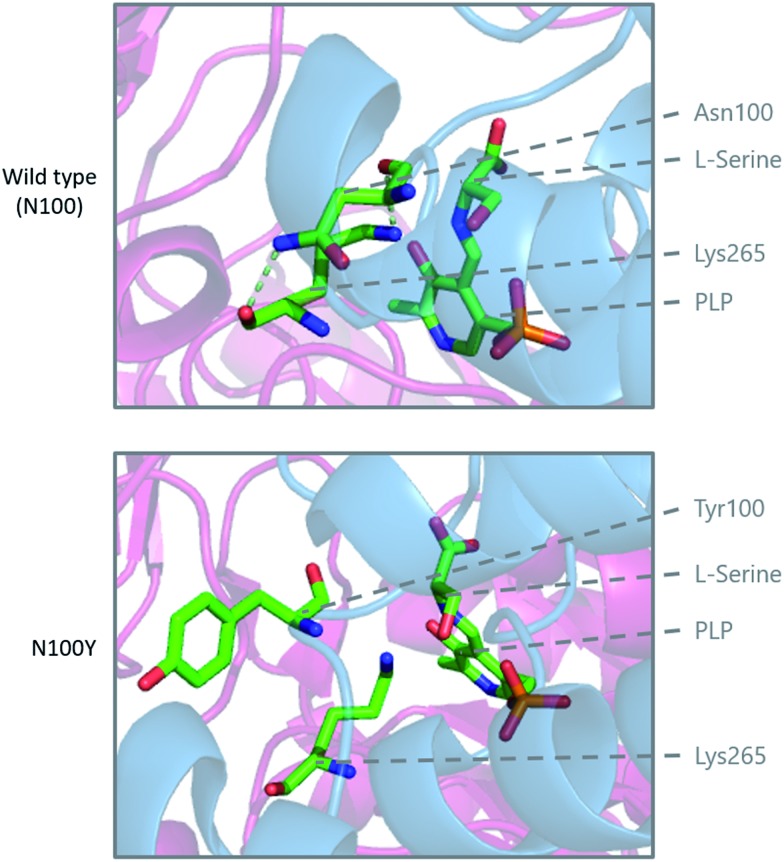
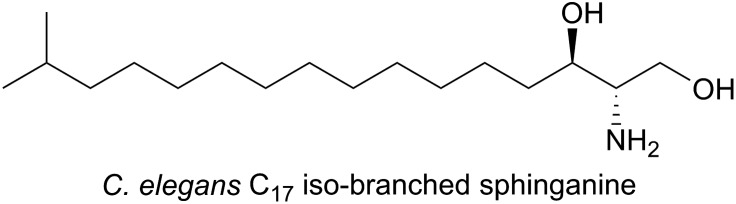
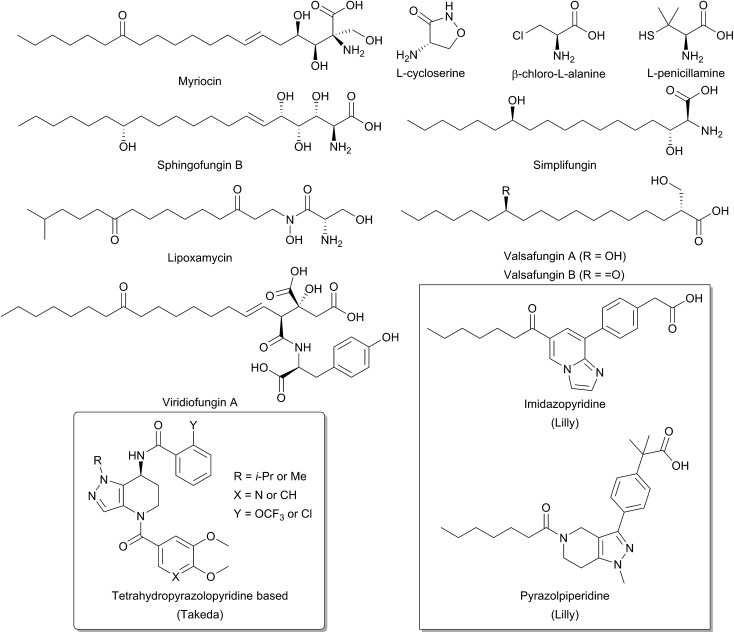
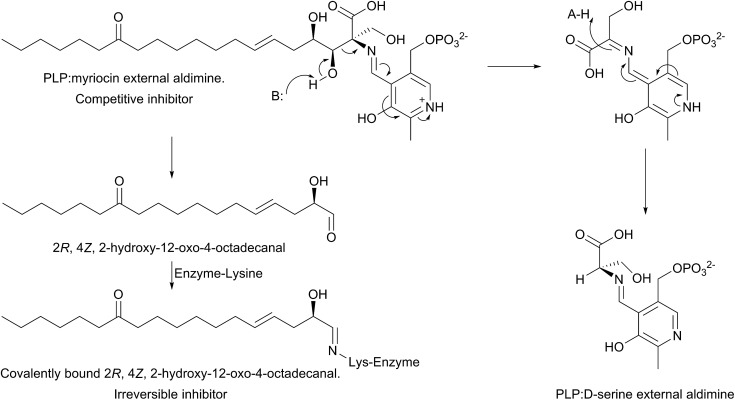
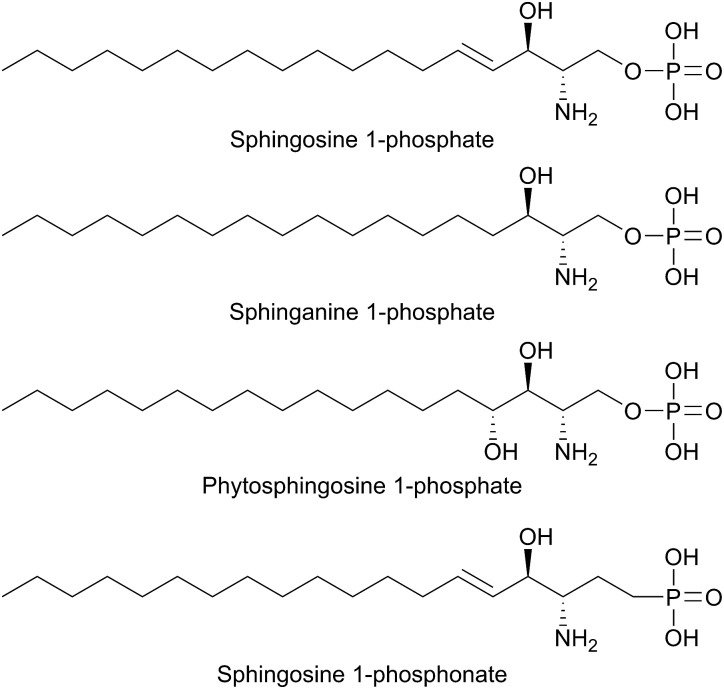
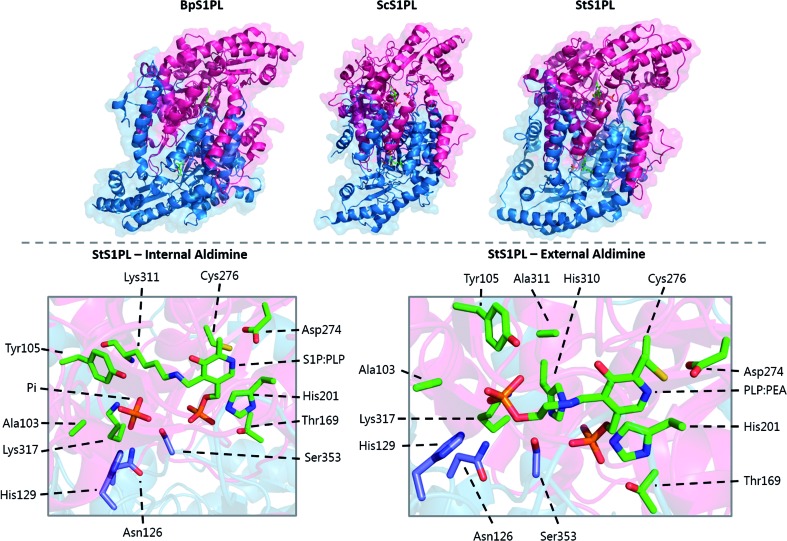
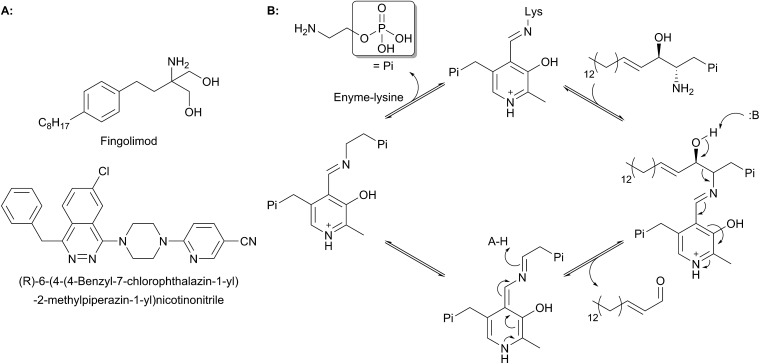



References
-
- Thudichum J. L. W., A Treatise on the Chemical Constitution of the Brain, 1884.
-
- Harayama T., Riezman H. Nat. Rev. Mol. Cell Biol. 2018;19:281–296. - PubMed
-
- Obeid L., Linardic C., Karolak L., Hannun Y. Science. 1993;259:1769–1771. - PubMed
-
- Okazaki T., Bell R. M., Hannun Y. A. J. Biol. Chem. 1989;264:19076–19080. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources