

Neuronatin regulates pancreatic β cell insulin content and secretion
- PMID: 29864031
- PMCID: PMC6063487
- DOI: 10.1172/JCI120115
Neuronatin regulates pancreatic β cell insulin content and secretion
Abstract
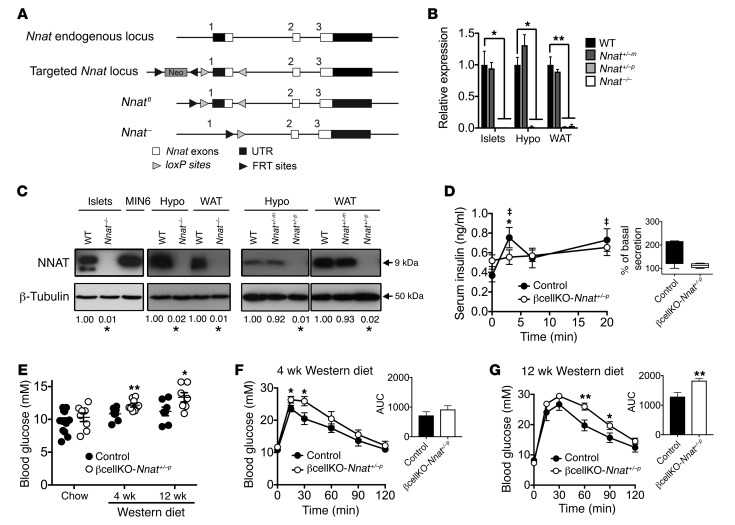
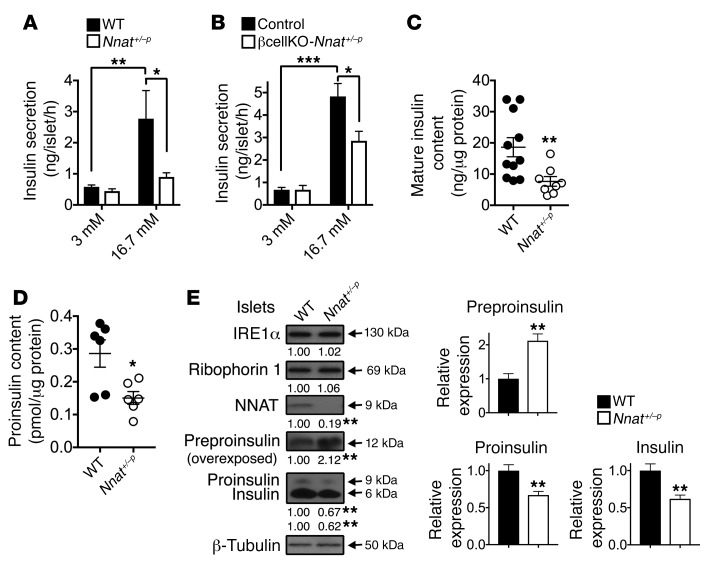
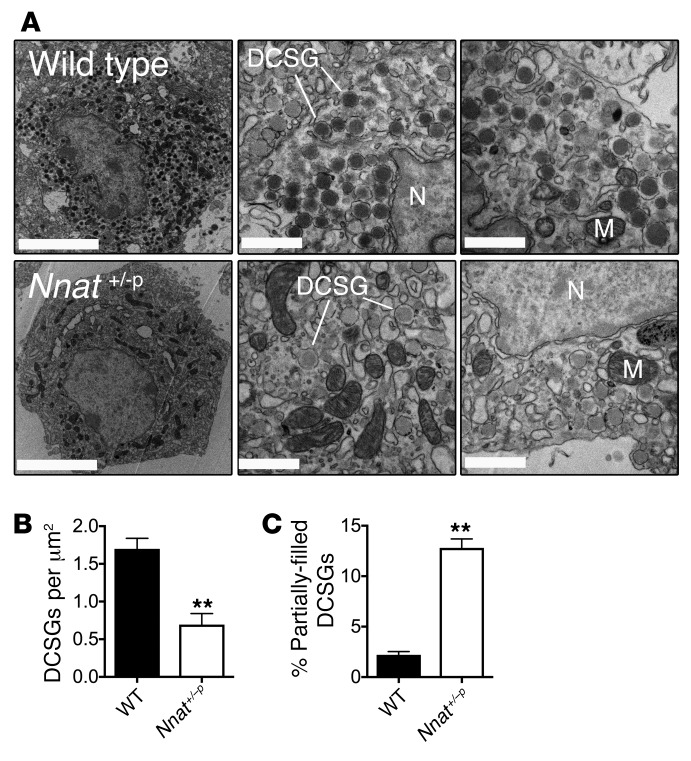
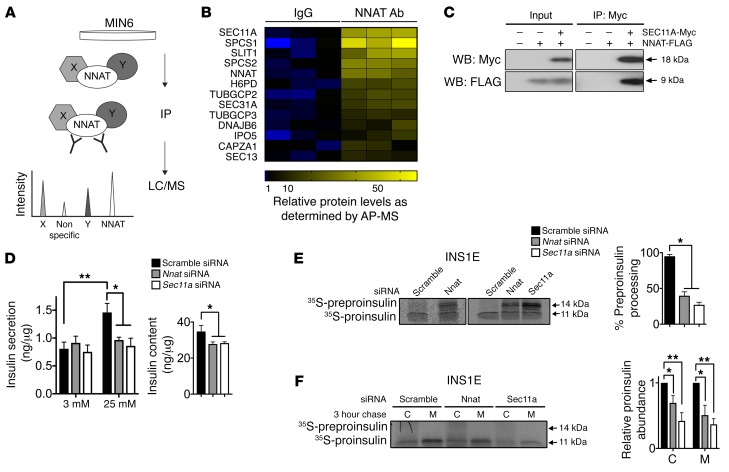
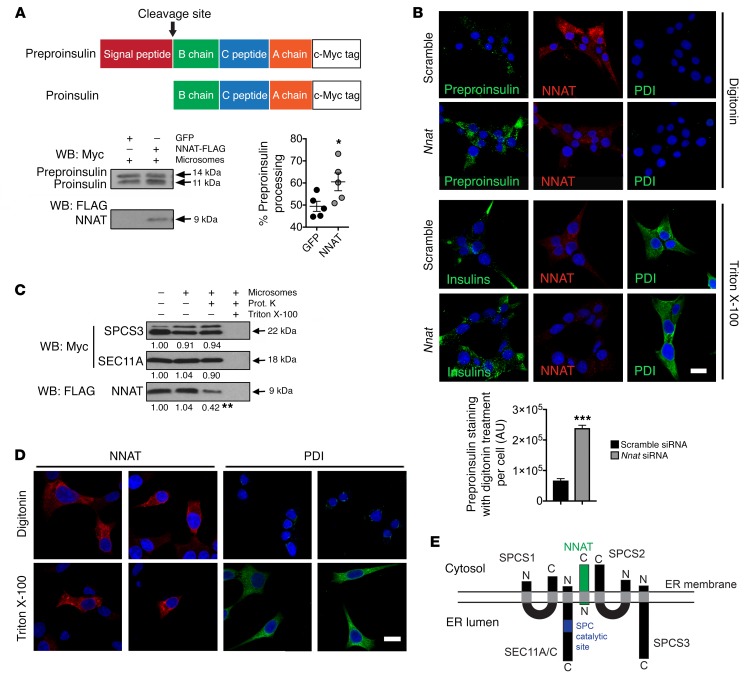
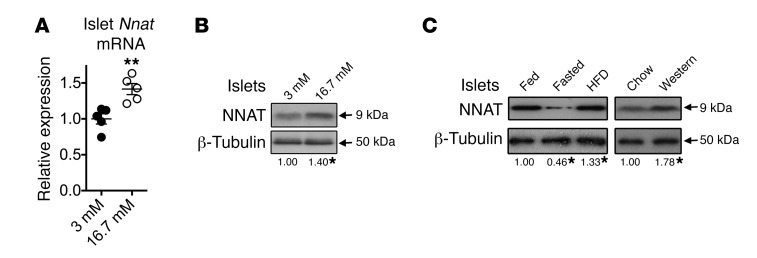
Neuronatin (Nnat) is an imprinted gene implicated in human obesity and widely expressed in neuroendocrine and metabolic tissues in a hormone- and nutrient-sensitive manner. However, its molecular and cellular functions and precise role in organismal physiology remain only partly defined. Here we demonstrate that mice lacking Nnat globally or specifically in β cells display impaired glucose-stimulated insulin secretion leading to defective glucose handling under conditions of nutrient excess. In contrast, we report no evidence for any feeding or body weight phenotypes in global Nnat-null mice. At the molecular level neuronatin augments insulin signal peptide cleavage by binding to the signal peptidase complex and facilitates translocation of the nascent preprohormone. Loss of neuronatin expression in β cells therefore reduces insulin content and blunts glucose-stimulated insulin secretion. Nnat expression, in turn, is glucose-regulated. This mechanism therefore represents a novel site of nutrient-sensitive control of β cell function and whole-animal glucose homeostasis. These data also suggest a potential wider role for Nnat in the regulation of metabolism through the modulation of peptide processing events.
Keywords: Beta cells; Cell Biology; Diabetes; Genetics; Insulin.
Conflict of interest statement
Figures
References
-
- Dou D, Joseph R. Cloning of human neuronatin gene and its localization to chromosome-20q 11.2-12: the deduced protein is a novel “proteolipid”. Brain Res. 1996;723(1–2):8–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MR/L020149/1/MRC_/Medical Research Council/United Kingdom
- MC_UP_1102/15/MRC_/Medical Research Council/United Kingdom
- MR/N00275X/1/MRC_/Medical Research Council/United Kingdom
- MR/N020472/1/MRC_/Medical Research Council/United Kingdom
- MR/K023667/1/MRC_/Medical Research Council/United Kingdom
- MR/M012646/1/MRC_/Medical Research Council/United Kingdom
- MR/L02036X/1/MRC_/Medical Research Council/United Kingdom
- MR/K001981/1/MRC_/Medical Research Council/United Kingdom
- FC001179/WT_/Wellcome Trust/United Kingdom
- MR/R010676/1/MRC_/Medical Research Council/United Kingdom
- MC_U120097114/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
