

Sox30 initiates transcription of haploid genes during late meiosis and spermiogenesis in mouse testes
- PMID: 29866902
- PMCID: PMC6053656
- DOI: 10.1242/dev.164855
Sox30 initiates transcription of haploid genes during late meiosis and spermiogenesis in mouse testes
Erratum in
-
Correction: Sox30 initiates transcription of haploid genes during late meiosis and spermiogenesis in mouse testes (doi:10.1242/dev.164855).Development. 2019 May 21;146(10):dev179978. doi: 10.1242/dev.179978. Development. 2019. PMID: 31113793 Free PMC article. No abstract available.
Abstract
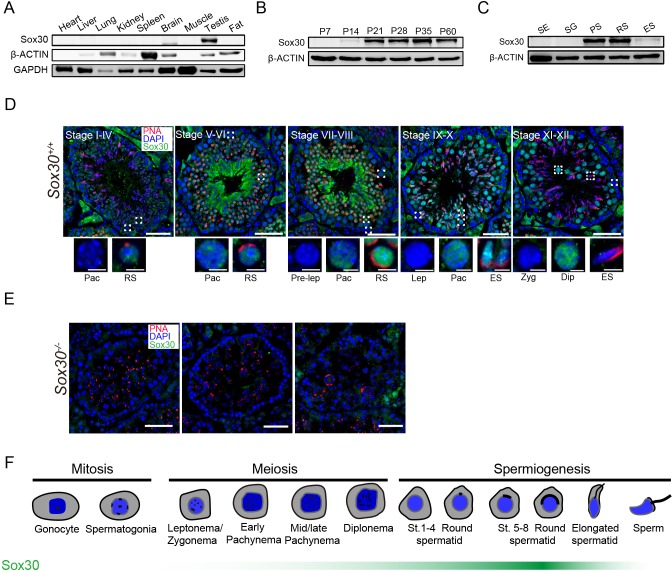
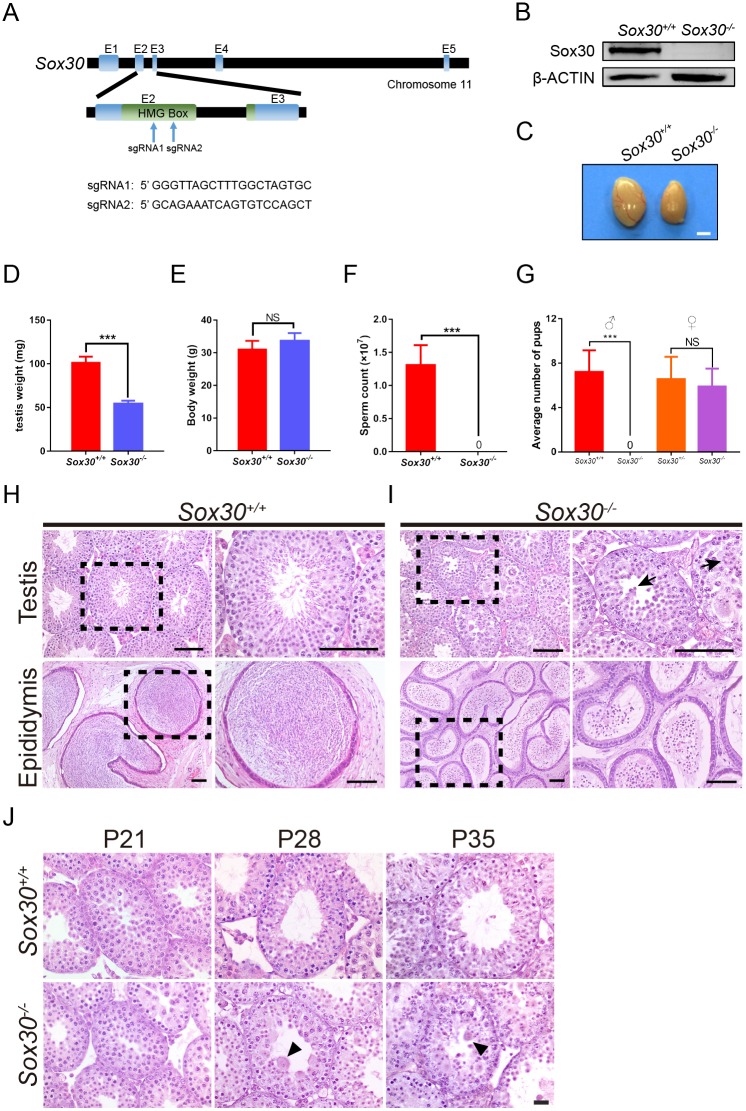
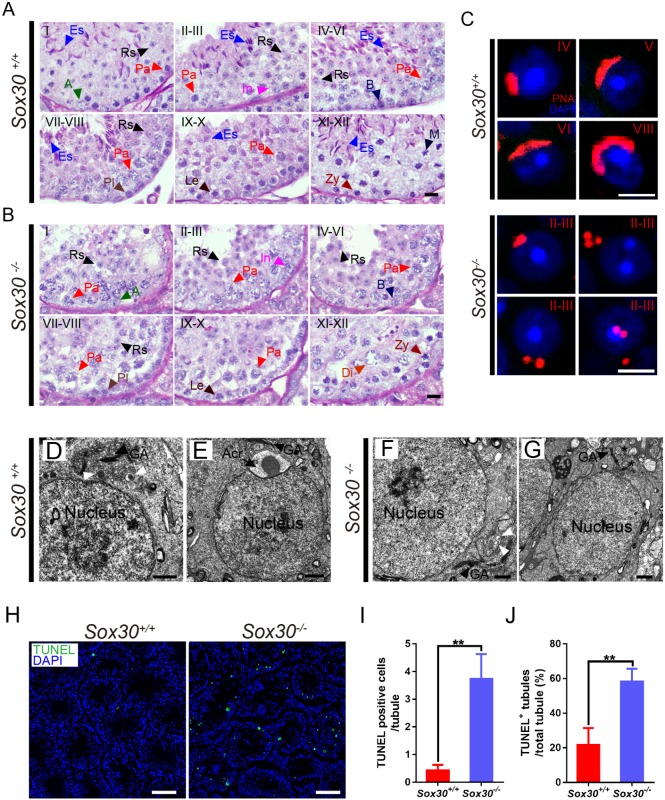
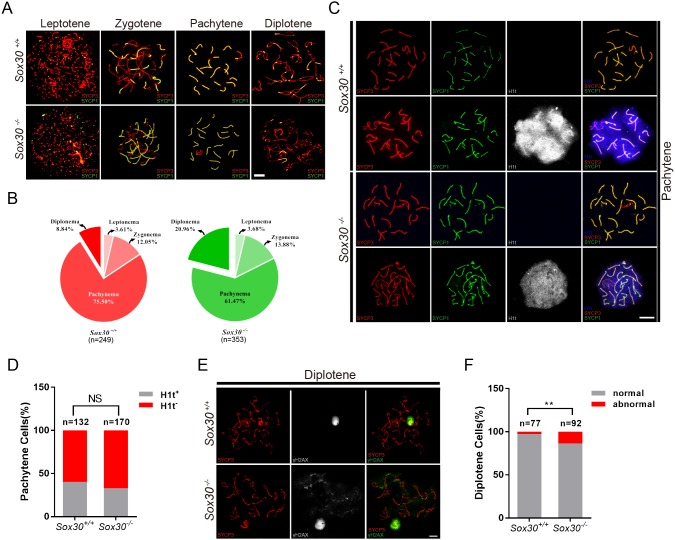
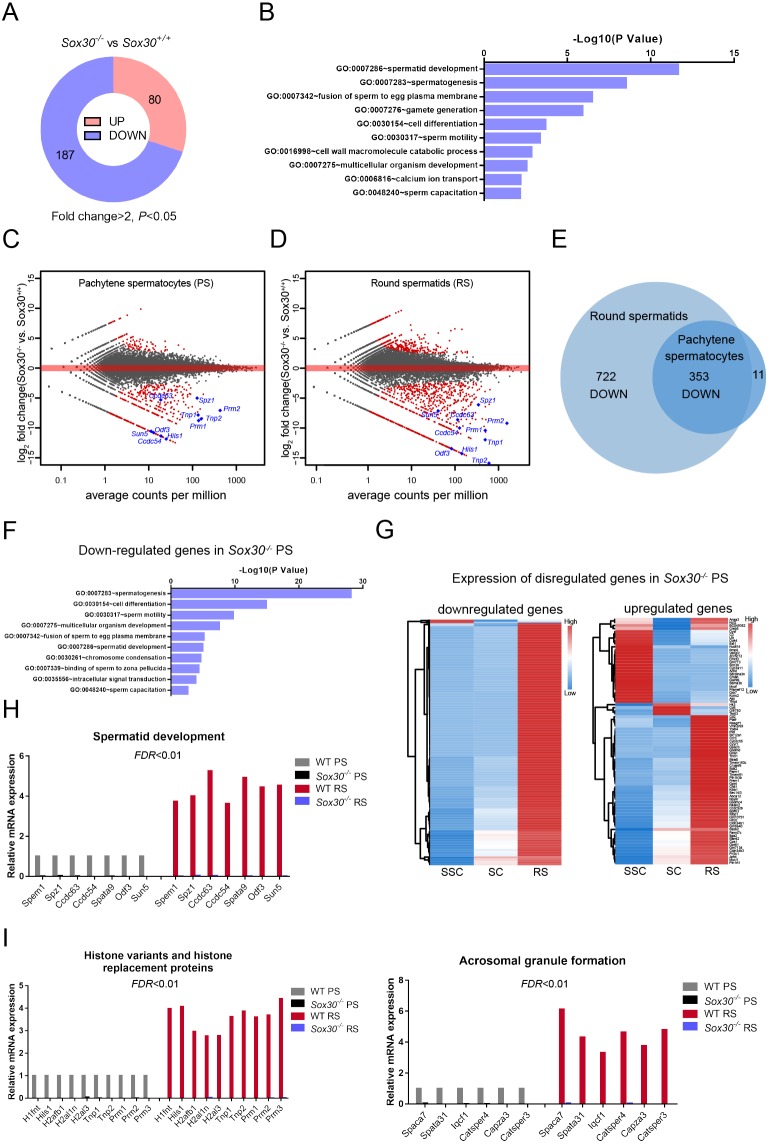
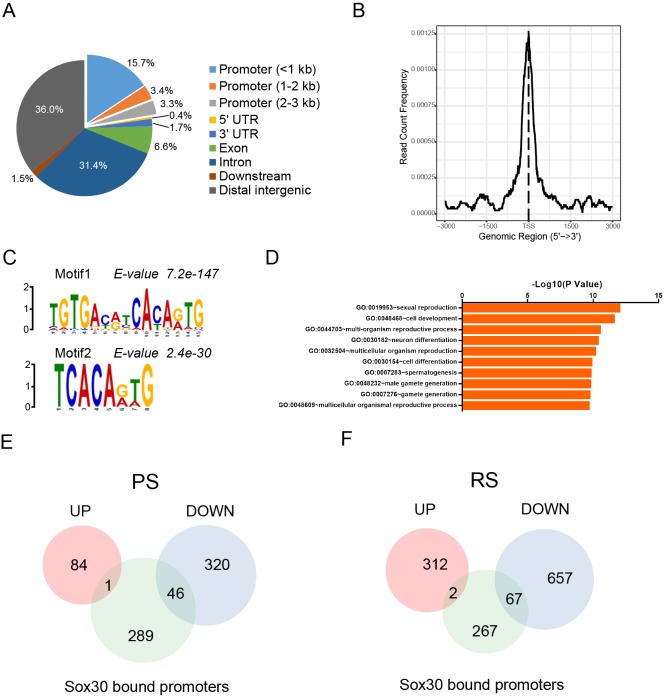
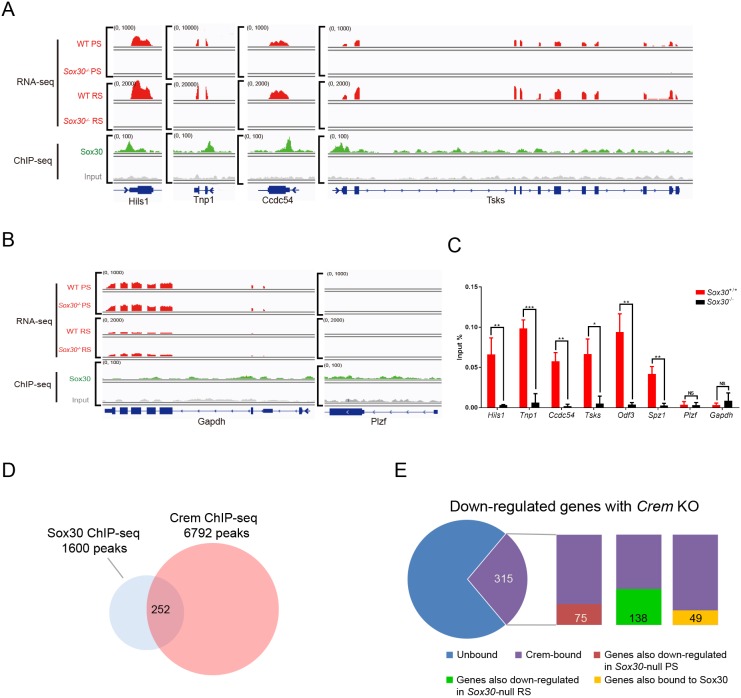
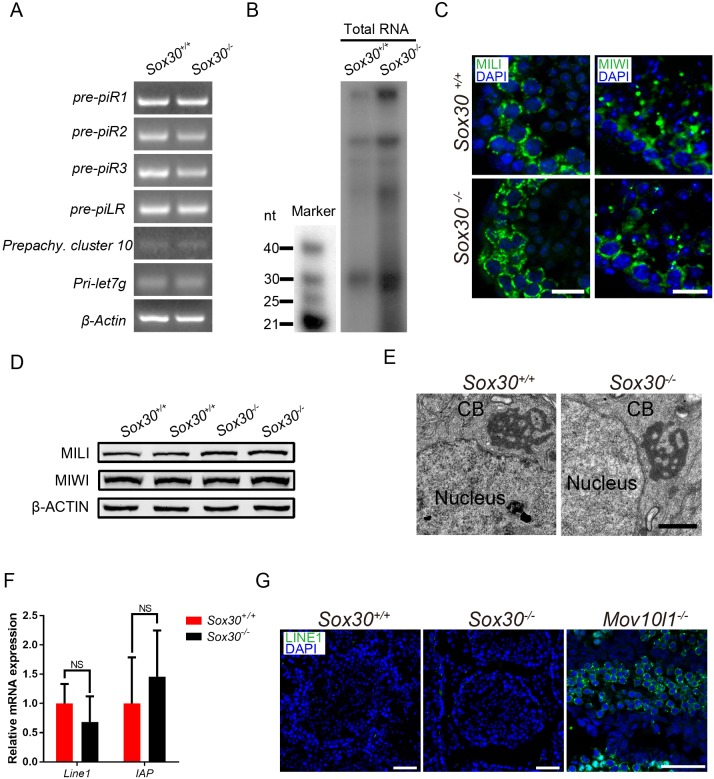
Transcription factors of the Sox protein family contain a DNA-binding HMG box and are key regulators of progenitor cell fate. Here, we report that expression of Sox30 is restricted to meiotic spermatocytes and postmeiotic haploids. Sox30 mutant males are sterile owing to spermiogenic arrest at the early round spermatid stage. Specifically, in the absence of Sox30, proacrosomic vesicles fail to form a single acrosomal organelle, and spermatids arrest at step 2-3. Although most Sox30 mutant spermatocytes progress through meiosis, accumulation of diplotene spermatocytes indicates a delayed or impaired transition from meiotic to postmeiotic stages. Transcriptome analysis of isolated stage-specific spermatogenic cells reveals that Sox30 controls a core postmeiotic gene expression program that initiates as early as the late meiotic cell stage. ChIP-seq analysis shows that Sox30 binds to specific DNA sequences in mouse testes, and its genomic occupancy correlates positively with expression of many postmeiotic genes including Tnp1, Hils1, Ccdc54 and Tsks These results define Sox30 as a crucial transcription factor that controls the transition from a late meiotic to a postmeiotic gene expression program and subsequent round spermatid development.
Keywords: Gene regulation; Male germ cell; Mouse; Sox30; Spermiogenesis.
© 2018. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
Similar articles
-
The transcription factor SOX30 is a key regulator of mouse spermiogenesis.Development. 2018 May 30;145(11):dev164723. doi: 10.1242/dev.164723. Development. 2018. PMID: 29848638
-
HDAC3 controls male fertility through enzyme-independent transcriptional regulation at the meiotic exit of spermatogenesis.Nucleic Acids Res. 2021 May 21;49(9):5106-5123. doi: 10.1093/nar/gkab313. Nucleic Acids Res. 2021. PMID: 33939832 Free PMC article.
-
SOX30 is required for male fertility in mice.Sci Rep. 2017 Dec 15;7(1):17619. doi: 10.1038/s41598-017-17854-5. Sci Rep. 2017. PMID: 29247201 Free PMC article.
-
A possible meiotic function of the peculiar patterns of gene expression in mammalian spermatogenic cells.Mech Dev. 2001 Aug;106(1-2):3-23. doi: 10.1016/s0925-4773(01)00413-0. Mech Dev. 2001. PMID: 11472831 Review.
-
[RNA-binding protein PTB in spermatogenesis: Progress in studies].Zhonghua Nan Ke Xue. 2016 Sep;22(9):856-860. Zhonghua Nan Ke Xue. 2016. PMID: 29071887 Review. Chinese.
Cited by
-
A 50-bp enhancer of the mouse acrosomal vesicle protein 1 gene activates round spermatid-specific transcription in vivo†.Biol Reprod. 2019 Oct 25;101(4):842-853. doi: 10.1093/biolre/ioz115. Biol Reprod. 2019. PMID: 31290539 Free PMC article.
-
SOX13 gene downregulation in peripheral blood mononuclear cells of patients with Klinefelter syndrome.Asian J Androl. 2021 Mar-Apr;23(2):157-162. doi: 10.4103/aja.aja_37_20. Asian J Androl. 2021. PMID: 33109779 Free PMC article.
-
The MYBL1/TCFL5 transcription network: two collaborative factors with central role in male meiosis.Biochem Soc Trans. 2023 Dec 20;51(6):2163-2172. doi: 10.1042/BST20231007. Biochem Soc Trans. 2023. PMID: 38015556 Free PMC article. Review.
-
A case of hermaphroditism in the gonochoristic sea urchin, Strongylocentrotus purpuratus, reveals key mechanisms of sex determination†.Biol Reprod. 2023 Jun 9;108(6):960-973. doi: 10.1093/biolre/ioad036. Biol Reprod. 2023. PMID: 36943312 Free PMC article.
-
Dnajb8, a target gene of SOX30, is dispensable for male fertility in mice.PeerJ. 2020 Dec 21;8:e10582. doi: 10.7717/peerj.10582. eCollection 2020. PeerJ. 2020. PMID: 33391882 Free PMC article.
References
-
- Barral S., Morozumi Y., Tanaka H., Montellier E., Govin J., de Dieuleveult M., Charbonnier G., Coute Y., Puthier D., Buchou T. et al. (2017). Histone variant H2A.L.2 guides transition protein-dependent protamine assembly in male germ cells. Mol. Cell 66, 89-101 e8. 10.1016/j.molcel.2017.02.025. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
