

Heterosis Is a Systemic Property Emerging From Non-linear Genotype-Phenotype Relationships: Evidence From in Vitro Genetics and Computer Simulations
- PMID: 29868111
- PMCID: PMC5968397
- DOI: 10.3389/fgene.2018.00159
Heterosis Is a Systemic Property Emerging From Non-linear Genotype-Phenotype Relationships: Evidence From in Vitro Genetics and Computer Simulations
Abstract
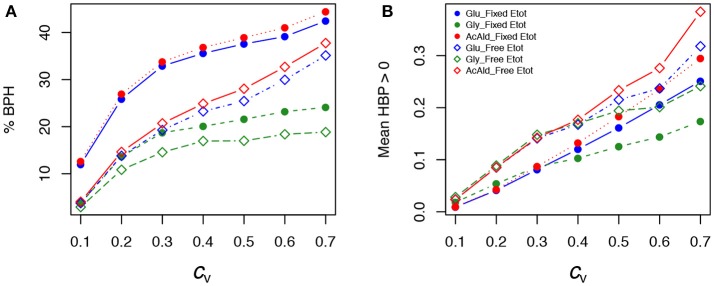
Heterosis, the superiority of hybrids over their parents for quantitative traits, represents a crucial issue in plant and animal breeding as well as evolutionary biology. Heterosis has given rise to countless genetic, genomic and molecular studies, but has rarely been investigated from the point of view of systems biology. We hypothesized that heterosis is an emergent property of living systems resulting from frequent concave relationships between genotypic variables and phenotypes, or between different phenotypic levels. We chose the enzyme-flux relationship as a model of the concave genotype-phenotype (GP) relationship, and showed that heterosis can be easily created in the laboratory. First, we reconstituted in vitro the upper part of glycolysis. We simulated genetic variability of enzyme activity by varying enzyme concentrations in test tubes. Mixing the content of "parental" tubes resulted in "hybrids," whose fluxes were compared to the parental fluxes. Frequent heterotic fluxes were observed, under conditions that were determined analytically and confirmed by computer simulation. Second, to test this model in a more realistic situation, we modeled the glycolysis/fermentation network in yeast by considering one input flux, glucose, and two output fluxes, glycerol and acetaldehyde. We simulated genetic variability by randomly drawing parental enzyme concentrations under various conditions, and computed the parental and hybrid fluxes using a system of differential equations. Again we found that a majority of hybrids exhibited positive heterosis for metabolic fluxes. Cases of negative heterosis were due to local convexity between certain enzyme concentrations and fluxes. In both approaches, heterosis was maximized when the parents were phenotypically close and when the distributions of parental enzyme concentrations were contrasted and constrained. These conclusions are not restricted to metabolic systems: they only depend on the concavity of the GP relationship, which is commonly observed at various levels of the phenotypic hierarchy, and could account for the pervasiveness of heterosis.
Keywords: enzyme variability; genotype-phenotype map; heterosis; mathematical modeling; metabolic network; non-linear processes.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures













References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases