

Toxoplasma gondii GRA8 induces ATP5A1-SIRT3-mediated mitochondrial metabolic resuscitation: a potential therapy for sepsis
- PMID: 29869623
- PMCID: PMC5898902
- DOI: 10.1038/emm.2017.308
Toxoplasma gondii GRA8 induces ATP5A1-SIRT3-mediated mitochondrial metabolic resuscitation: a potential therapy for sepsis
Abstract
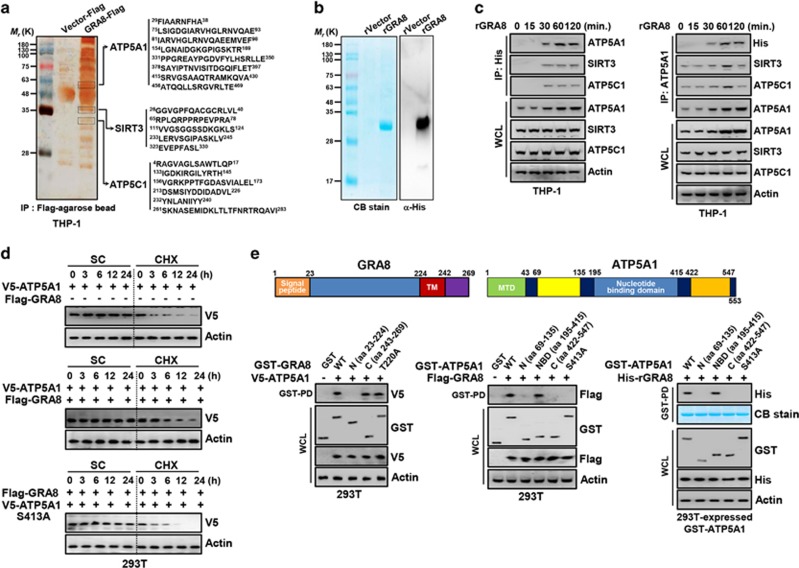
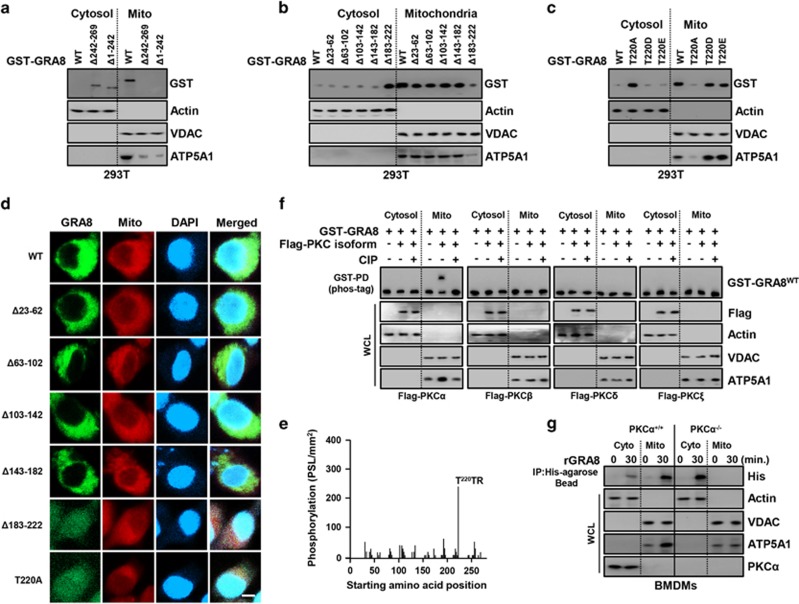
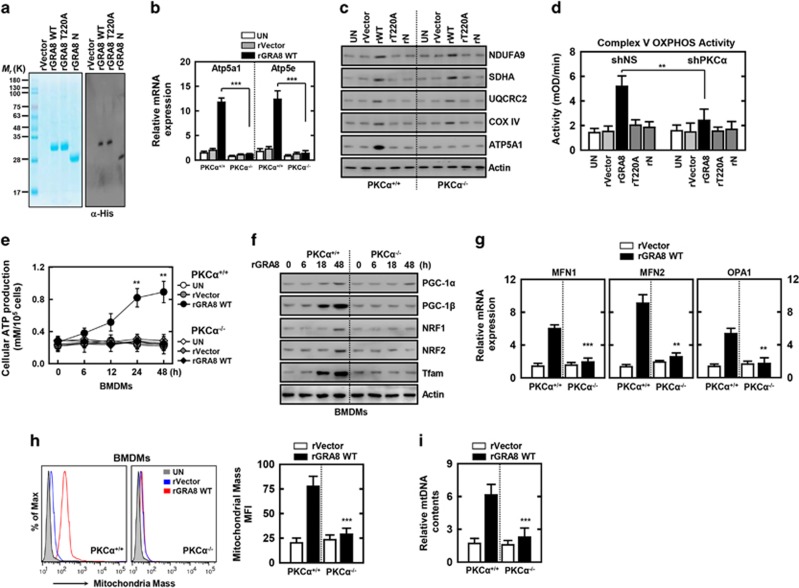
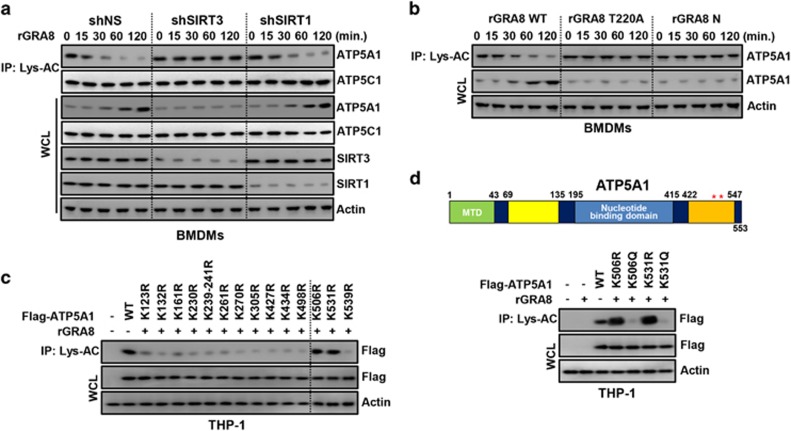
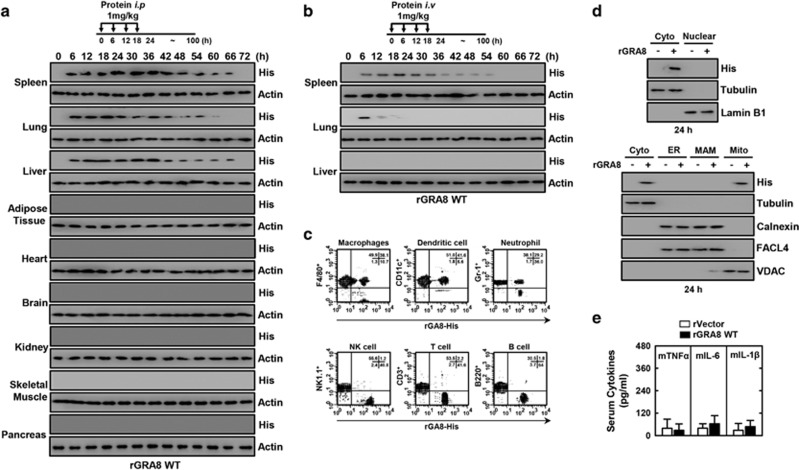
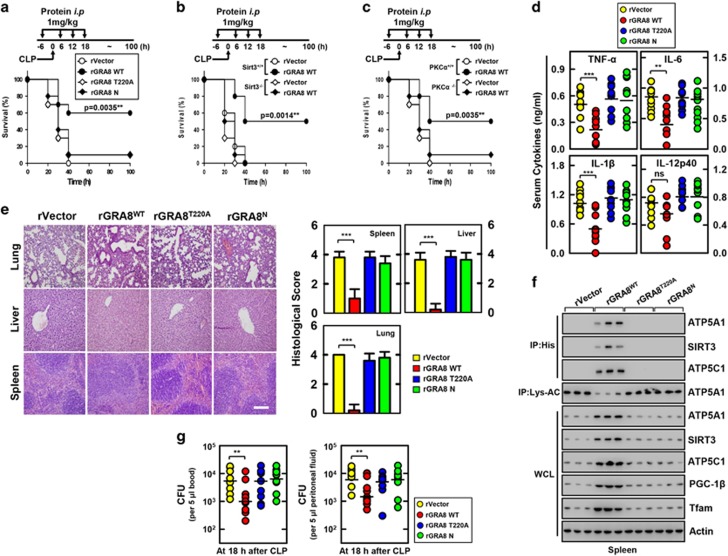
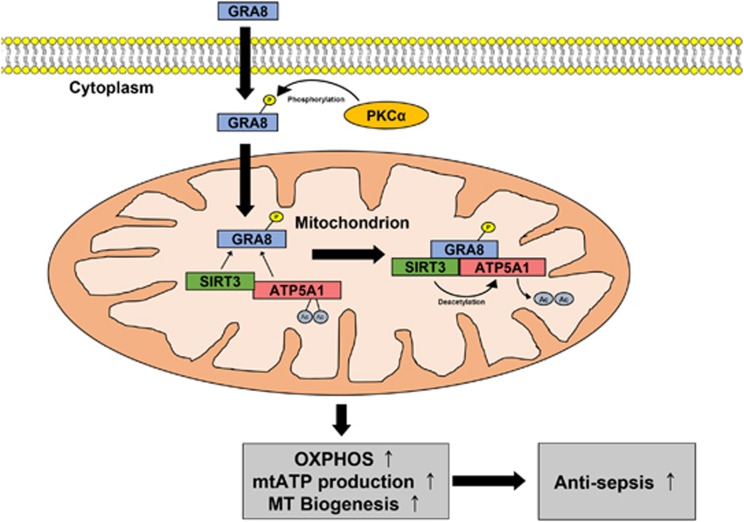
The intracellular parasite Toxoplasma gondii has unique dense granule antigens (GRAs) that are crucial for host infection. Emerging evidence suggests that GRA8 of T. gondii is a promising serodiagnostic marker in toxoplasmosis. However, little is known about the intracellular regulatory mechanisms involved in GRA8-induced host responses. We found that GRA8 interacts with host proteins involved in mitochondria activation and might be useful as a therapeutic strategy for sepsis. Here, we show that protein kinase-Cα (PKCα)-mediated phosphorylation of T. gondii GRA8 (Thr220) is required for mitochondrial trafficking and regulates the interaction of C terminal of GRA8 with nucleotide binding domain of ATP5A1. Furthermore, GRA8 interacts with SIRT3 in mitochondria, facilitating ATP5A1 deacetylation (K506 and K531), adenosine triphosphate production and subsequent anti-septic activity in vivo. Taken together, these results demonstrate a new anti-sepsis therapeutic strategy using T. gondii GRA8-induced mitochondrial metabolic resuscitation. This strategy represents an urgently needed paradigm shift for therapeutic intervention.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Koethe M, Pott S, Ludewig M, Bangoura B, Zoller B, Daugschies A et al. Prevalence of specific IgG-antibodies against Toxoplasma gondii in domestic turkeys determined by kinetic ELISA based on recombinant GRA7 and GRA8. Vet Parasitol 2011; 180: 179–190. - PubMed
-
- Costa JG, Dure AB. Immunochemical evaluation of two Toxoplasma gondii GRA8 sequences to detect acute toxoplasmosis infection. Microb Pathog 2016; 100: 229–236. - PubMed
-
- Kim YR, Hwang J, Koh HJ, Jang K, Lee JD, Choi J et al. The targeted delivery of the c-Src peptide complexed with schizophyllan to macrophages inhibits polymicrobial sepsis and ulcerative colitis in mice. Biomaterials 2016; 89: 1–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
