

Temporal profiling of redox-dependent heterogeneity in single cells
- PMID: 29869985
- PMCID: PMC6023615
- DOI: 10.7554/eLife.37623
Temporal profiling of redox-dependent heterogeneity in single cells
Abstract
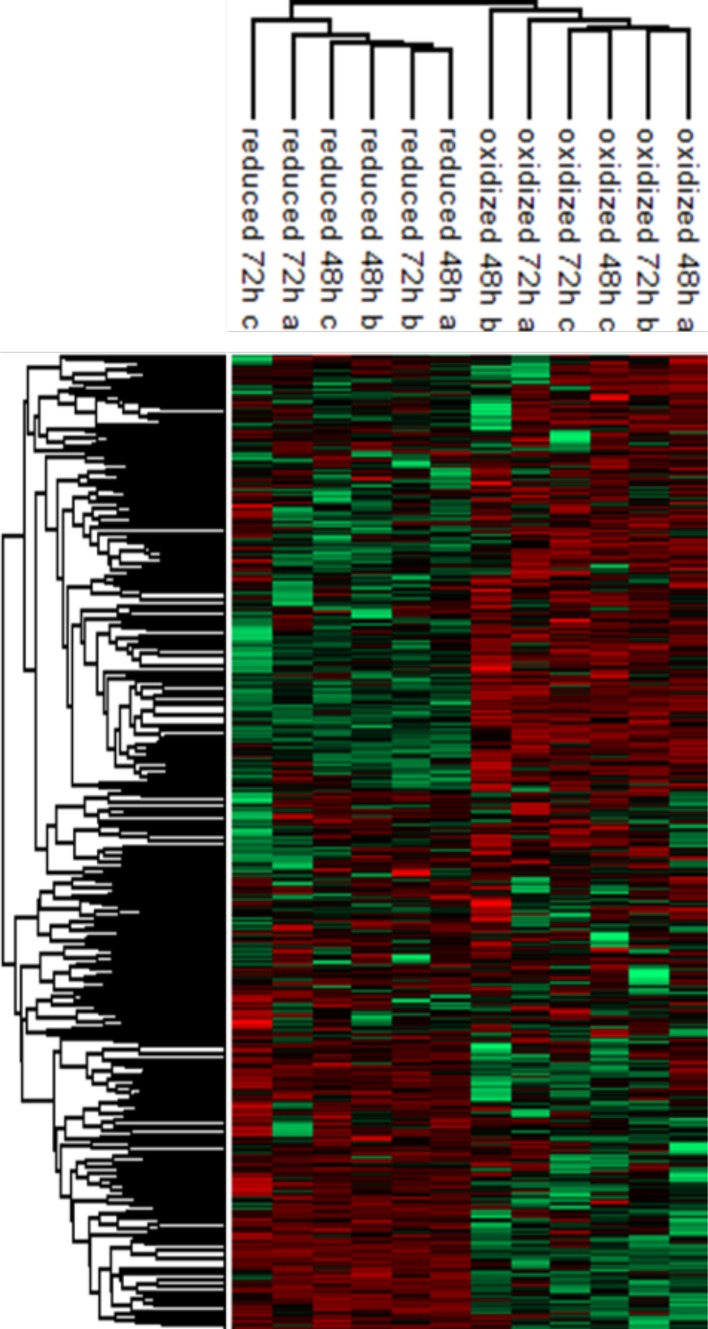
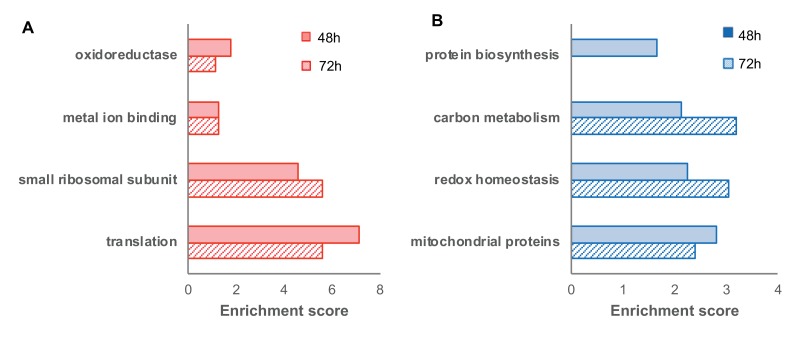
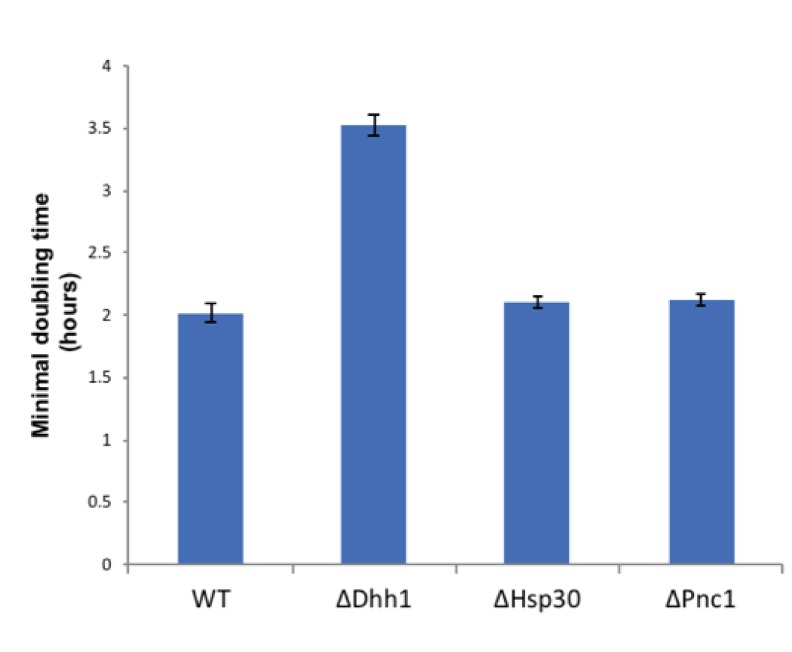
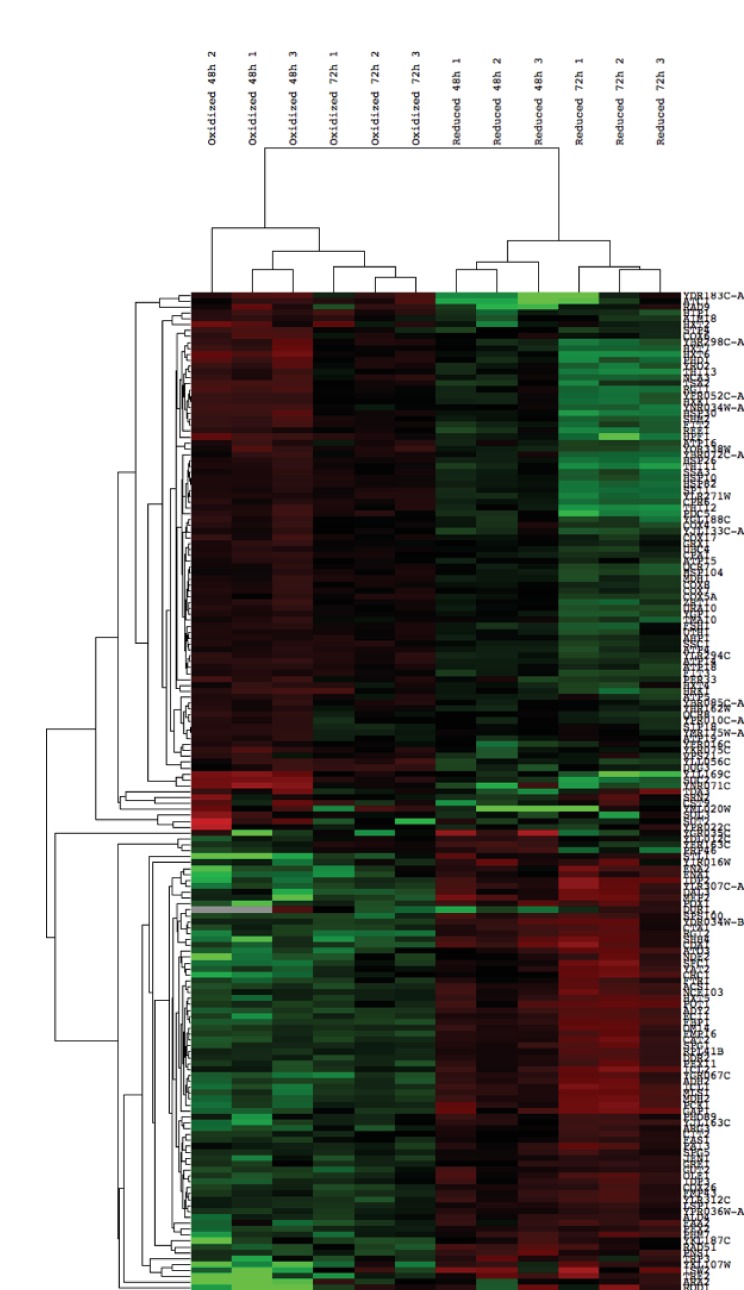
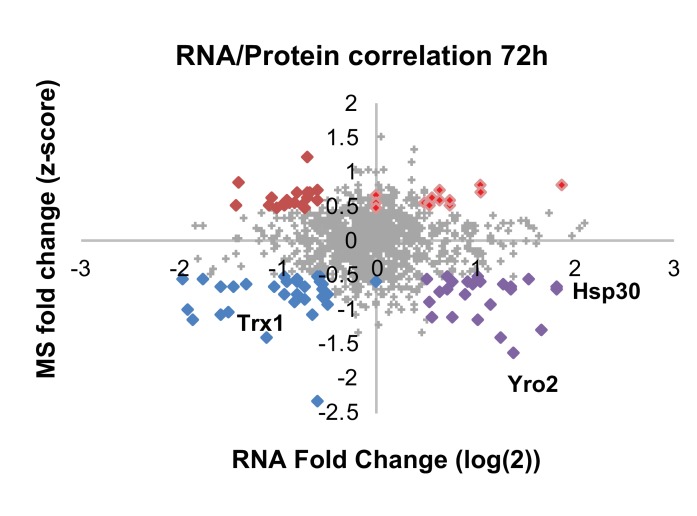
Cellular redox status affects diverse cellular functions, including proliferation, protein homeostasis, and aging. Thus, individual differences in redox status can give rise to distinct sub-populations even among cells with identical genetic backgrounds. Here, we have created a novel methodology to track redox status at single cell resolution using the redox-sensitive probe Grx1-roGFP2. Our method allows identification and sorting of sub-populations with different oxidation levels in either the cytosol, mitochondria or peroxisomes. Using this approach, we defined a redox-dependent heterogeneity of yeast cells and characterized growth, as well as proteomic and transcriptomic profiles of distinctive redox subpopulations. We report that, starting in late logarithmic growth, cells of the same age have a bi-modal distribution of oxidation status. A comparative proteomic analysis between these populations identified three key proteins, Hsp30, Dhh1, and Pnc1, which affect basal oxidation levels and may serve as first line of defense proteins in redox homeostasis.
Keywords: S. cerevisiae; biochemistry; cell biology; chemical biology; redox biology; redox sensors; roGFP; yeast.
© 2018, Radzinski et al.
Conflict of interest statement
MR, RF, OY, WB, NS, JG, SI, YG, ST, YN, TR, NF, MS, DR No competing interests declared
Figures

















References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
