

Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp
- PMID: 29870563
- PMCID: PMC5988331
- DOI: 10.1371/journal.pone.0198694
Stress cross-response of the antioxidative system promoted by superimposed drought and cold conditions in Coffea spp
Abstract
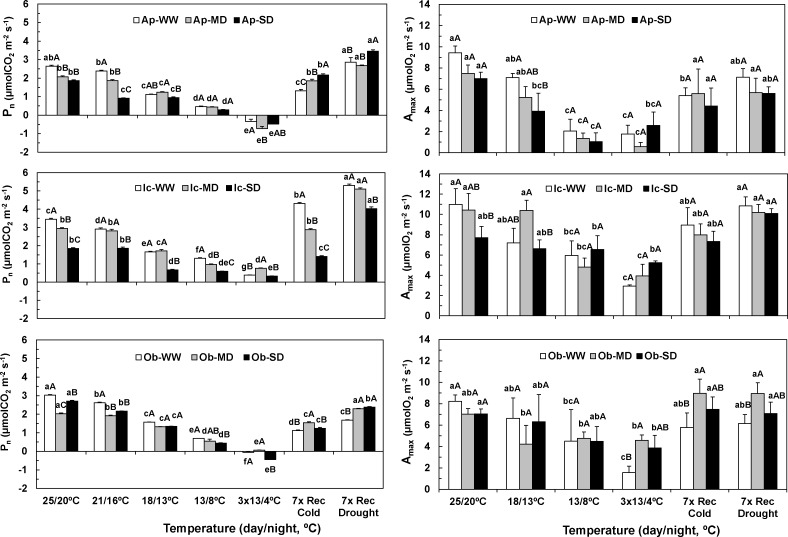
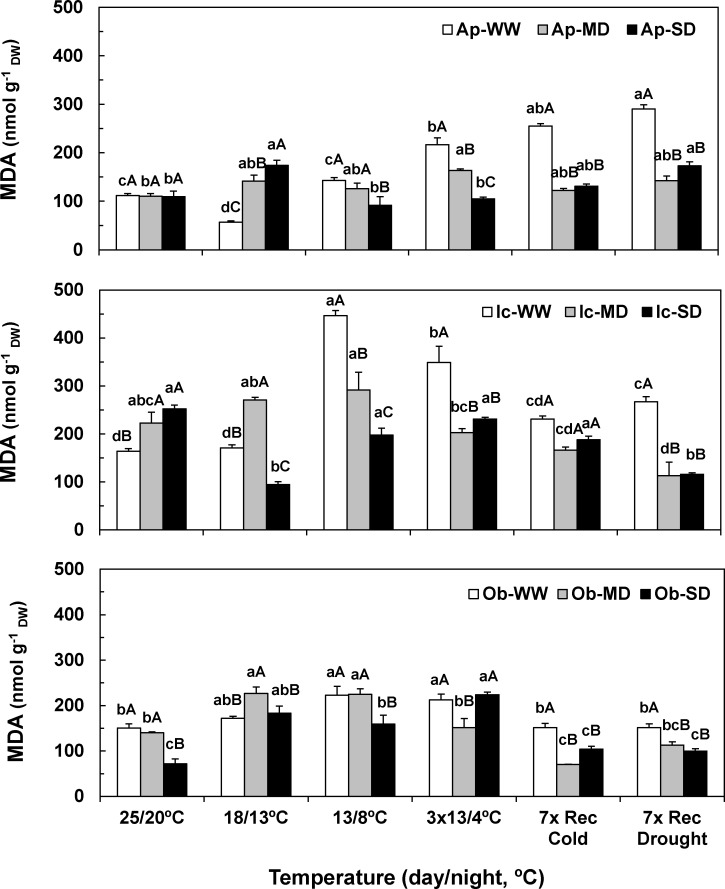
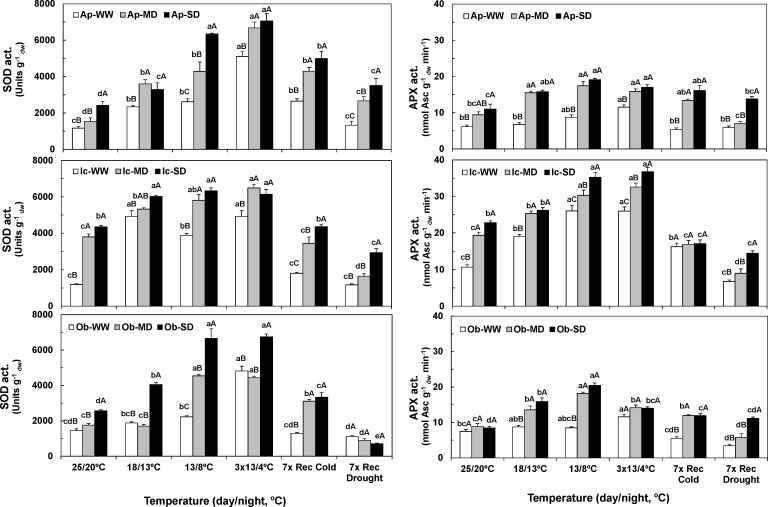
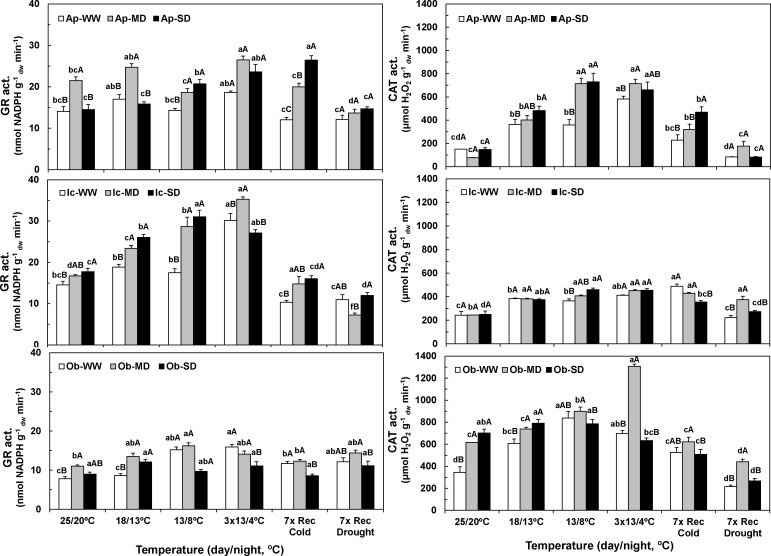
The understanding of acclimation strategies to low temperature and water availability is decisive to ensure coffee crop sustainability, since these environmental conditions determine the suitability of cultivation areas. In this context, the impacts of single and combined exposure to drought and cold were evaluated in three genotypes of the two major cropped species, Coffea arabica cv. Icatu, Coffea canephora cv. Apoatã, and the hybrid Obatã. Crucial traits of plant resilience to environmental stresses have been examined: photosynthesis, lipoperoxidation and the antioxidant response. Drought and/or cold promoted leaf dehydration, which was accompanied by stomatal and mesophyll limitations that impaired leaf C-assimilation in all genotypes. However, Icatu showed a lower impact upon stress exposure and a faster and complete photosynthetic recovery. Although lipoperoxidation was increased by drought (Icatu) and cold (all genotypes), it was greatly reduced by stress interaction, especially in Icatu. In fact, although the antioxidative system was reinforced under single drought and cold exposure (e.g., activity of enzymes as Cu,Zn-superoxide dismutase, ascorbate peroxidase, APX, glutathione reductase and catalase, CAT), the stronger increases were observed upon the simultaneous exposure to both stresses, which was accompanied with a transcriptional response of some genes, namely related to APX. Complementary, non-enzyme antioxidant molecules were promoted mostly by cold and the stress interaction, including α-tocopherol (in C. arabica plants), ascorbate (ASC), zeaxanthin, and phenolic compounds (all genotypes). In general, drought promoted antioxidant enzymes activity, whereas cold enhanced the synthesis of both enzyme and non-enzyme antioxidants, the latter likely related to a higher need of antioxidative capability when enzyme reactions were probably quite repressed by low temperature. Icatu showed the wider antioxidative capability, with the triggering of all studied antioxidative molecules by drought (except CAT), cold, and, particularly, stress interaction (except ASC), revealing a clear stress cross-tolerance. This justified the lower impacts on membrane lipoperoxidation and photosynthetic capacity under stress interaction conditions, related to a better ROS control. These findings are also relevant to coffee water management, showing that watering in the cold season should be largely avoided.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
-
- Wang W, Vinocur B, Altman A. Plant responses to drought, salinity and extreme temperatures: towards genetic engineering for stress tolerance. Planta 2003; 218: 1–14. doi: 10.1007/s00425-003-1105-5 - DOI - PubMed
-
- Way DA, Oren R, Kroner Y. The space-time continuum: the effects of elevated CO2 and temperature on trees and the importance of scaling. Plant Cell Environ. 2015; 38: 991–1007. doi: 10.1111/pce.12527 - DOI - PubMed
-
- Rodrigues WP, Martins MQ, Fortunato AS, Rodrigues AP, Semedo JN, Simões-Costa MC, et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Global Change Biol. 2016; 22: 415–431. doi: 10.1111/gcb.13088 - DOI - PubMed
-
- Des Marais DL, Lasky JR, Verslues PE, Chang TZ, Juenger TE. Interactive effects of water limitation and elevated temperature on the physiology, development and fitness of diverse accessions of Brachypodium distachyon. New Phytol. 2017; 214: 132–144. doi: 10.1111/nph.14316 - DOI - PubMed
-
- Fujita M, Fujita Y, Noutosh Y, Takahashi F, Narusaka Y, Yamaguchi-Shinozaki K, Shinozaki K. Crosstalk between abiotic and biotic stress responses: a current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006; 9: 436–442. doi: 10.1016/j.pbi.2006.05.014 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
