

Bicarbonate directly modulates activity of chemosensitive neurons in the retrotrapezoid nucleus
- PMID: 29873079
- PMCID: PMC6117565
- DOI: 10.1113/JP276104
Bicarbonate directly modulates activity of chemosensitive neurons in the retrotrapezoid nucleus
Abstract
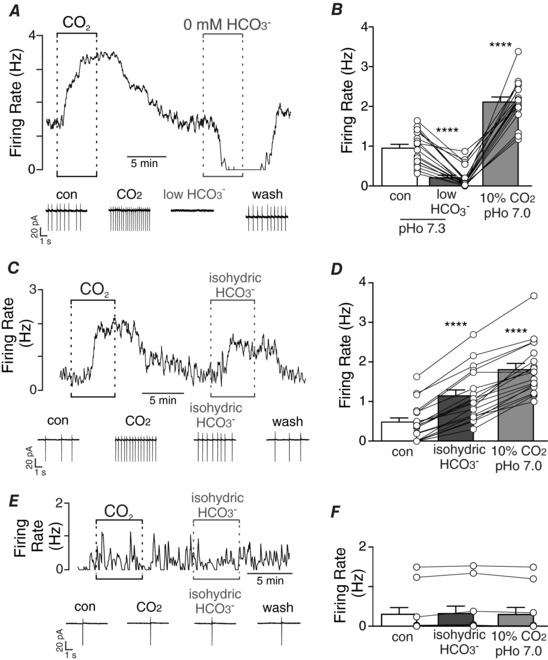
Key points: Changes in CO2 result in corresponding changes in both H+ and HCO3- and despite evidence that HCO3- can function as an independent signalling molecule, there is little evidence suggesting HCO3- contributes to respiratory chemoreception. We show that HCO3- directly activates chemosensitive retrotrapezoid nucleus (RTN) neurons. Identifying all relevant signalling molecules is essential for understanding how chemoreceptors function, and because HCO3- and H+ are buffered by separate cellular mechanisms, having the ability to sense both modalities adds additional information regarding changes in CO2 that are not necessarily reflected by pH alone. HCO3- may be particularly important for regulating activity of RTN chemoreceptors during sustained intracellular acidifications when TASK-2 channels, which appear to be the sole intracellular pH sensor, are minimally active.
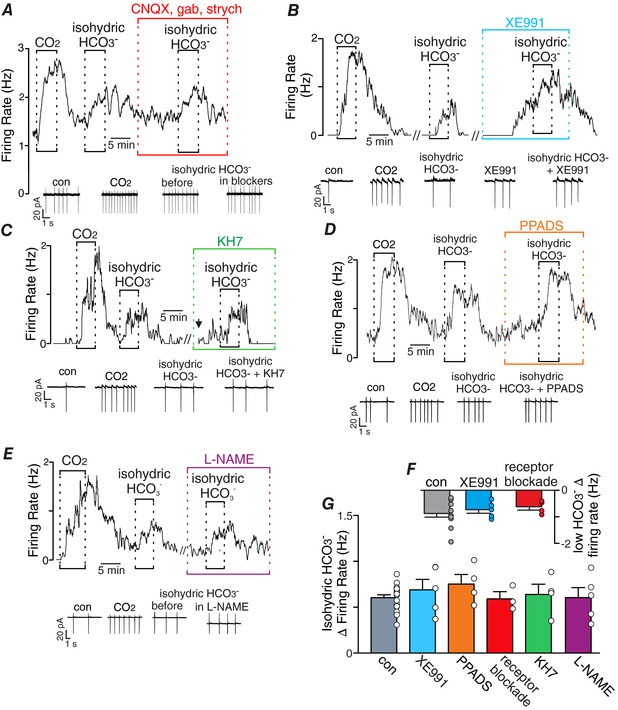
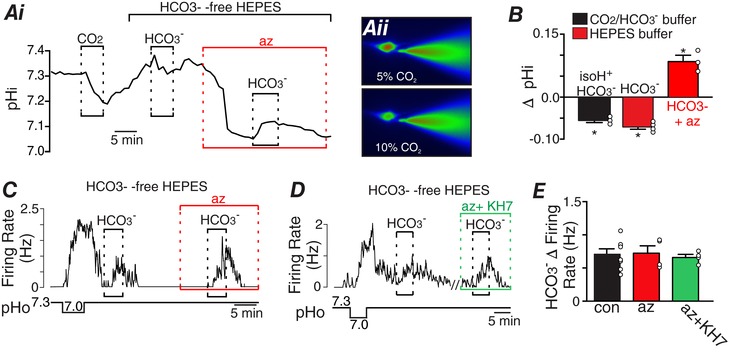
Abstract: Central chemoreception is the mechanism by which the brain regulates breathing in response to changes in tissue CO2 /H+ . The retrotrapezoid nucleus (RTN) is an important site of respiratory chemoreception. Mechanisms underlying RTN chemoreception involve H+ -mediated activation of chemosensitive neurons and CO2 /H+ -evoked ATP-purinergic signalling by local astrocytes, which activates chemosensitive neurons directly and indirectly by maintaining vascular tone when CO2 /H+ levels are high. Although changes in CO2 result in corresponding changes in both H+ and HCO3- and despite evidence that HCO3- can function as an independent signalling molecule, there is little evidence suggesting HCO3- contributes to respiratory chemoreception. Therefore, the goal of this study was to determine whether HCO3- regulates activity of chemosensitive RTN neurons independent of pH. Cell-attached recordings were used to monitor activity of chemosensitive RTN neurons in brainstem slices (300 μm thick) isolated from rat pups (postnatal days 7-11) during exposure to low or high concentrations of HCO3- . In a subset of experiments, we also included 2',7'-bis(2carboxyethyl)-5-(and 6)-carboxyfluorescein (BCECF) in the internal solution to measure pHi under each experimental condition. We found that HCO3- activates chemosensitive RTN neurons by mechanisms independent of intracellular or extracellular pH, glutamate, GABA, glycine or purinergic signalling, soluble adenylyl cyclase activity, nitric oxide or KCNQ channels. These results establish HCO3- as a novel independent modulator of chemoreceptor activity, and because the levels of HCO3- along with H+ are buffered by independent cellular mechanisms, these results suggest HCO3- chemoreception adds additional information regarding changes in CO2 that are not necessarily reflected by pH.
Keywords: HCO3; brain slice; chemoreception; pH-independent; pHi.
© 2018 The Authors. The Journal of Physiology © 2018 The Physiological Society.
Figures
Comment in
-
Breaking down CO2 : bicarbonate the other side of central chemosensitivity in retrotrapezoid nucleus neurons.J Physiol. 2018 Sep;596(17):3827-3828. doi: 10.1113/JP276558. Epub 2018 Jul 30. J Physiol. 2018. PMID: 29978487 Free PMC article. No abstract available.
References
-
- Bonnet U & Bingmann D (1993). GABA‐responses of CA3 neurones at epileptogenic threshold concentrations of convulsants. Neuroreport 4, 715–718. - PubMed
-
- Chen Y, Cann MJ, Litvin TN, Iourgenko V, Sinclair ML, Levin LR & Buck J (2000). Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 289, 625–628. - PubMed
-
- Chesler M (2003). Regulation and modulation of pH in the brain. Physiol Rev 83, 1183–1221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous