

The Melatonin Analog IQM316 May Induce Adult Hippocampal Neurogenesis and Preserve Recognition Memories in Mice
- PMID: 29873251
- PMCID: PMC6038050
- DOI: 10.1177/0963689717721217
The Melatonin Analog IQM316 May Induce Adult Hippocampal Neurogenesis and Preserve Recognition Memories in Mice
Abstract
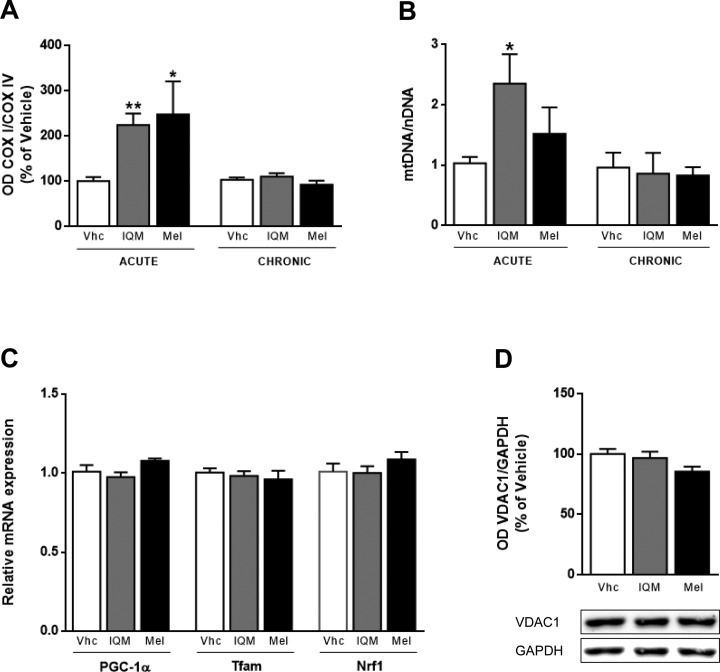
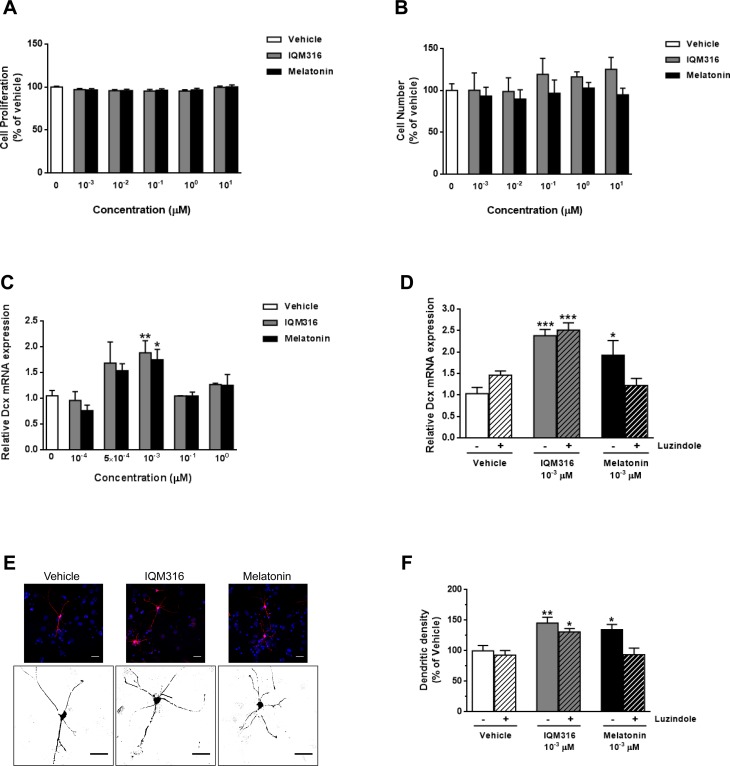
Neurogenesis in the adult hippocampus is a unique process in neurobiology that requires functional integration of newly generated neurons, which may disrupt existing hippocampal network connections and consequently loss of established memories. As neurodegenerative diseases characterized by abnormal neurogenesis and memory dysfunctions are increasing, the identification of new anti-aging drugs is required. In adult mice, we found that melatonin, a well-established neurogenic hormone, and the melatonin analog 2-(2-(5-methoxy-1 H-indol-3-yl)ethyl)-5-methyl-1,3,4-oxadiazole (IQM316) were able to induce hippocampal neurogenesis, measured by neuronal nuclei (NeuN) and 5-bromo-2'-deoxyuridine (BrdU) labeling. More importantly, only IQM316 administration was able to induce hippocampal neurogenesis while preserving previously acquired memories, assessed with object recognition tests. In vitro studies with embryonic neural stem cells replicated the finding that both melatonin and IQM316 induce direct differentiation of neural precursors without altering their proliferative activity. Furthermore, IQM316 induces differentiation through a mechanism that is not dependent of melatonergic receptors (MTRs), since the MTR antagonist luzindole could not block the IQM316-induced effects. We also found that IQM316 and melatonin modulate mitochondrial DNA copy number and oxidative phosphorylation proteins, while maintaining mitochondrial function as measured by respiratory assays and enzymatic activity. These results uncover a novel pharmacological agent that may be capable of inducing adult hippocampal neurogenesis at a healthy and sustainable rate that preserves recognition memories.
Keywords: adult neurogenesis; long-term memory; melatonin; neural stem cells.
Conflict of interest statement
Figures
Similar articles
-
Chronic administration of a melatonin membrane receptor antagonist, luzindole, affects hippocampal neurogenesis without changes in hopelessness-like behavior in adult mice.Neuropharmacology. 2016 Apr;103:211-21. doi: 10.1016/j.neuropharm.2015.11.030. Epub 2015 Dec 12. Neuropharmacology. 2016. PMID: 26686389
-
Melatonin attenuates methamphetamine-induced inhibition of neurogenesis in the adult mouse hippocampus: An in vivo study.Neurosci Lett. 2015 Oct 8;606:209-14. doi: 10.1016/j.neulet.2015.09.011. Epub 2015 Sep 11. Neurosci Lett. 2015. PMID: 26366944
-
Melatonin modulates cell survival of new neurons in the hippocampus of adult mice.Neuropsychopharmacology. 2009 Aug;34(9):2180-91. doi: 10.1038/npp.2009.46. Epub 2009 May 6. Neuropsychopharmacology. 2009. PMID: 19421166
-
Benefits of the Neurogenic Potential of Melatonin for Treating Neurological and Neuropsychiatric Disorders.Int J Mol Sci. 2023 Mar 2;24(5):4803. doi: 10.3390/ijms24054803. Int J Mol Sci. 2023. PMID: 36902233 Free PMC article. Review.
-
Effects of melatonin and its analogues on neural stem cells.Mol Cell Endocrinol. 2016 Jan 15;420:169-79. doi: 10.1016/j.mce.2015.10.012. Epub 2015 Oct 21. Mol Cell Endocrinol. 2016. PMID: 26499395 Review.
Cited by
-
Adult Neurogenesis under Control of the Circadian System.Cells. 2022 Feb 22;11(5):764. doi: 10.3390/cells11050764. Cells. 2022. PMID: 35269386 Free PMC article. Review.
-
Melatonin Enhances Neural Differentiation of Adipose-Derived Mesenchymal Stem Cells.Int J Mol Sci. 2024 Apr 30;25(9):4891. doi: 10.3390/ijms25094891. Int J Mol Sci. 2024. PMID: 38732109 Free PMC article.
-
Protective Effects of Melatonin on Neurogenesis Impairment in Neurological Disorders and Its Relevant Molecular Mechanisms.Int J Mol Sci. 2020 Aug 6;21(16):5645. doi: 10.3390/ijms21165645. Int J Mol Sci. 2020. PMID: 32781737 Free PMC article. Review.
-
Is Adult Hippocampal Neurogenesis Really Relevant for the Treatment of Psychiatric Disorders?Curr Neuropharmacol. 2021;19(10):1640-1660. doi: 10.2174/1570159X18666200818194948. Curr Neuropharmacol. 2021. PMID: 32811415 Free PMC article. Review.
-
Recent Neurotherapeutic Strategies to Promote Healthy Brain Aging: Are we there yet?Aging Dis. 2022 Feb 1;13(1):175-214. doi: 10.14336/AD.2021.0705. eCollection 2022 Feb. Aging Dis. 2022. PMID: 35111369 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
