

The Role of Network Architecture in Collagen Mechanics
- PMID: 29874616
- PMCID: PMC6129505
- DOI: 10.1016/j.bpj.2018.04.043
The Role of Network Architecture in Collagen Mechanics
Abstract
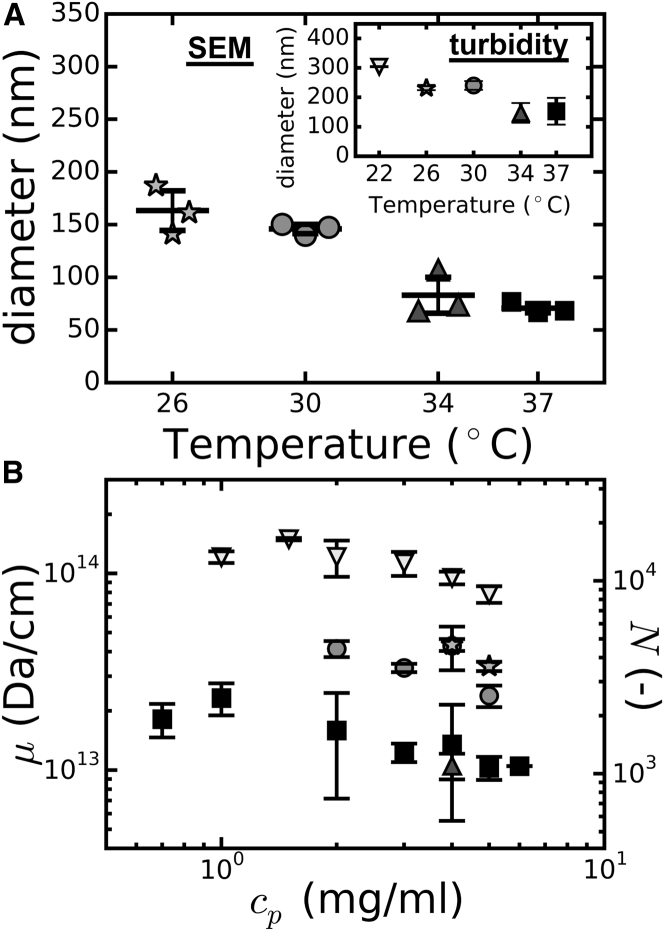
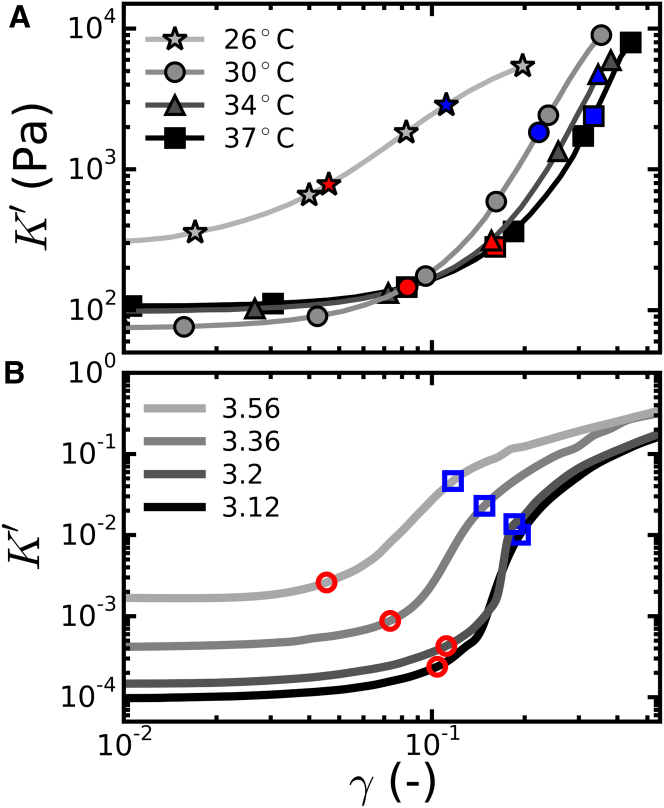
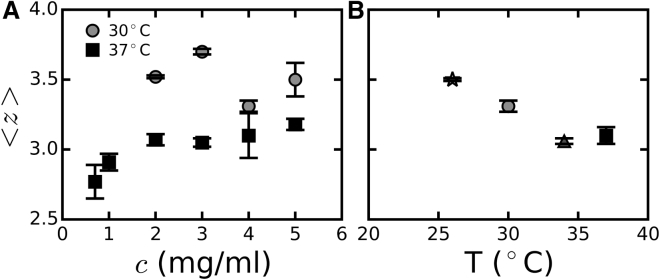
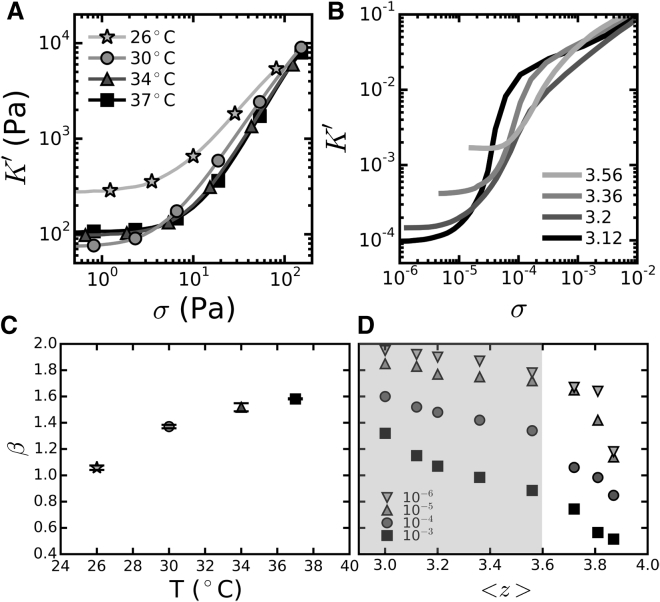
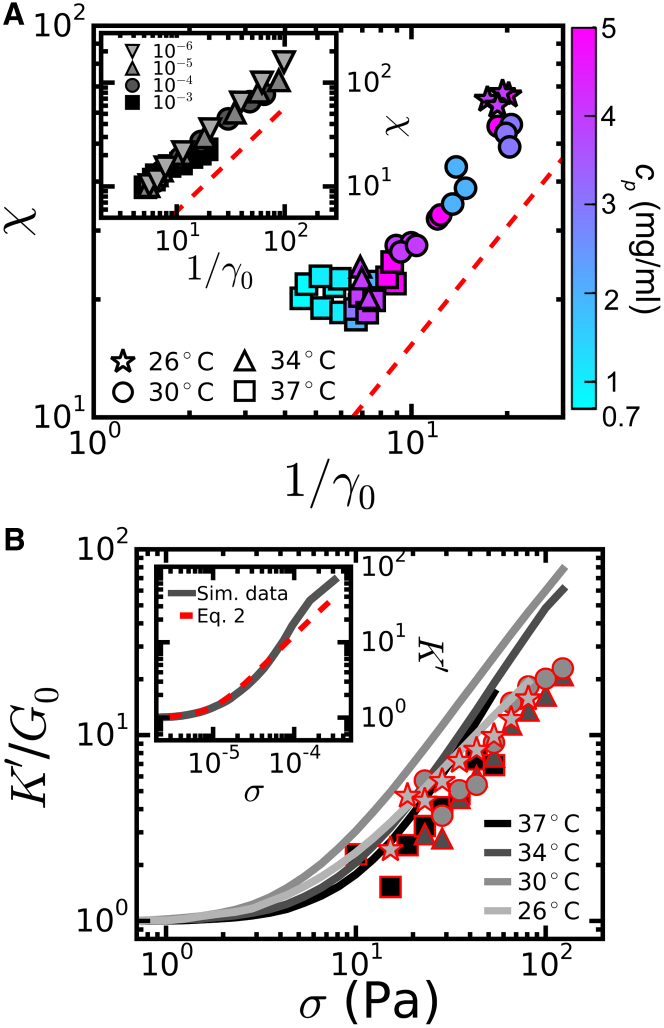
Collagen forms fibrous networks that reinforce tissues and provide an extracellular matrix for cells. These networks exhibit remarkable strain-stiffening properties that tailor the mechanical functions of tissues and regulate cell behavior. Recent models explain this nonlinear behavior as an intrinsic feature of disordered networks of stiff fibers. Here, we experimentally validate this theoretical framework by measuring the elastic properties of collagen networks over a wide range of self-assembly conditions. We show that the model allows us to quantitatively relate both the linear and nonlinear elastic behavior of collagen networks to their underlying architecture. Specifically, we identify the local coordination number (or connectivity) 〈z〉 as a key architectural parameter that governs the elastic response of collagen. The network elastic response reveals that 〈z〉 decreases from 3.5 to 3 as the polymerization temperature is raised from 26 to 37°C while being weakly dependent on concentration. We furthermore infer a Young's modulus of 1.1 MPa for the collagen fibrils from the linear modulus. Scanning electron microscopy confirms that 〈z〉 is between three and four but is unable to detect the subtle changes in 〈z〉 with polymerization conditions that rheology is sensitive to. Finally, we show that, consistent with the model, the initial stress-stiffening response of collagen networks is controlled by the negative normal stress that builds up under shear. Our work provides a predictive framework to facilitate future studies of the regulatory effect of extracellular matrix molecules on collagen mechanics. Moreover, our findings can aid mechanobiological studies of wound healing, fibrosis, and cancer metastasis, which require collagen matrices with tunable mechanical properties.
Copyright © 2018 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Fratzl P., editor. Collagen Structure and Mechanics. Springer Science+Business Media; Berlin, Germany: 2008.
-
- Kjaer M., Langberg H., Magnusson S.P. From mechanical loading to collagen synthesis, structural changes and function in human tendon. Scand. J. Med. Sci. Sports. 2009;19:500–510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
