

Assembly and maintenance of GABAergic and Glycinergic circuits in the mammalian nervous system
- PMID: 29875009
- PMCID: PMC5991458
- DOI: 10.1186/s13064-018-0109-6
Assembly and maintenance of GABAergic and Glycinergic circuits in the mammalian nervous system
Abstract
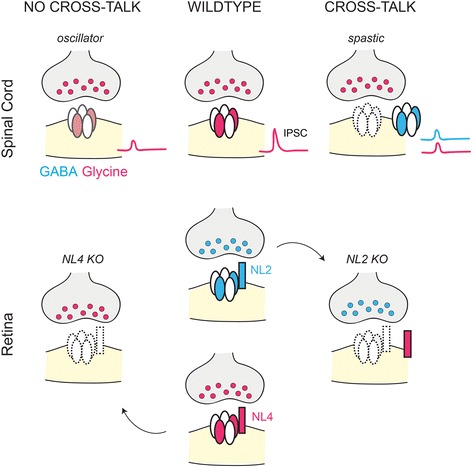
Inhibition in the central nervous systems (CNS) is mediated by two neurotransmitters: gamma-aminobutyric acid (GABA) and glycine. Inhibitory synapses are generally GABAergic or glycinergic, although there are synapses that co-release both neurotransmitter types. Compared to excitatory circuits, much less is known about the cellular and molecular mechanisms that regulate synaptic partner selection and wiring patterns of inhibitory circuits. Recent work, however, has begun to fill this gap in knowledge, providing deeper insight into whether GABAergic and glycinergic circuit assembly and maintenance rely on common or distinct mechanisms. Here we summarize and contrast the developmental mechanisms that regulate the selection of synaptic partners, and that promote the formation, refinement, maturation and maintenance of GABAergic and glycinergic synapses and their respective wiring patterns. We highlight how some parts of the CNS demonstrate developmental changes in the type of inhibitory transmitter or receptor composition at their inhibitory synapses. We also consider how perturbation of the development or maintenance of one type of inhibitory connection affects other inhibitory synapse types in the same circuit. Mechanistic insight into the development and maintenance of GABAergic and glycinergic inputs, and inputs that co-release both these neurotransmitters could help formulate comprehensive therapeutic strategies for treating disorders of synaptic inhibition.
Keywords: Circuit refinement; Inhibition; Synapse formation; Synapse maturation; Synaptic targeting.
Conflict of interest statement
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Figures
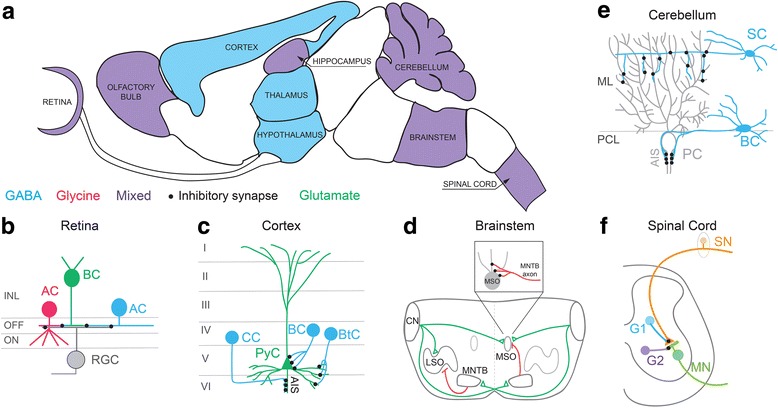
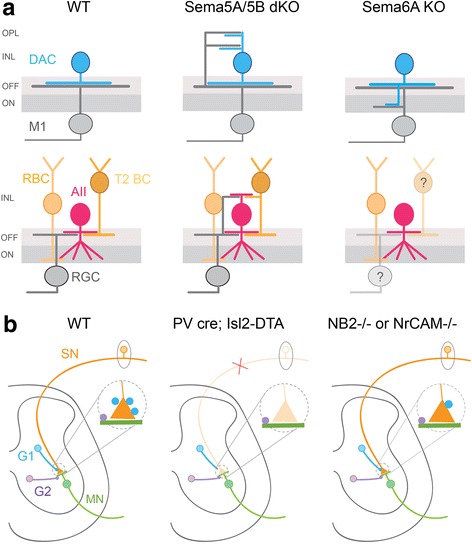
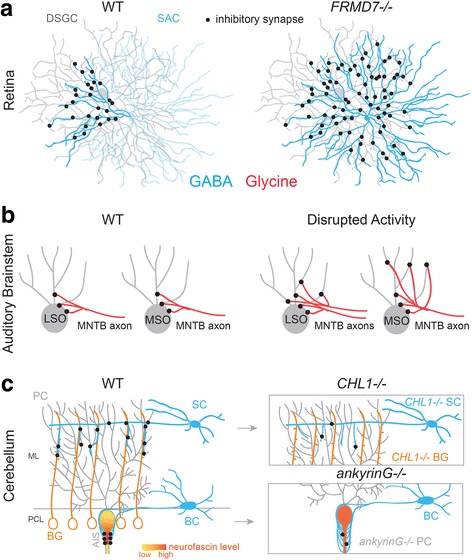
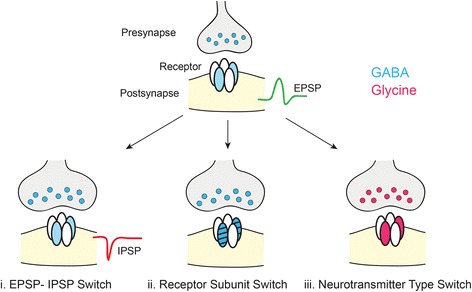
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
