

Flexible Sensorimotor Computations through Rapid Reconfiguration of Cortical Dynamics
- PMID: 29879384
- PMCID: PMC6009852
- DOI: 10.1016/j.neuron.2018.05.020
Flexible Sensorimotor Computations through Rapid Reconfiguration of Cortical Dynamics
Abstract
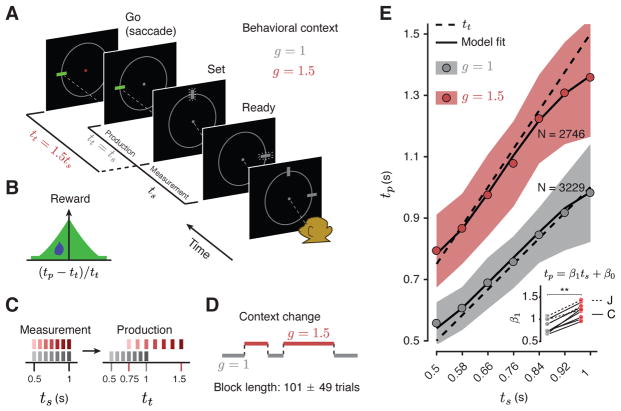
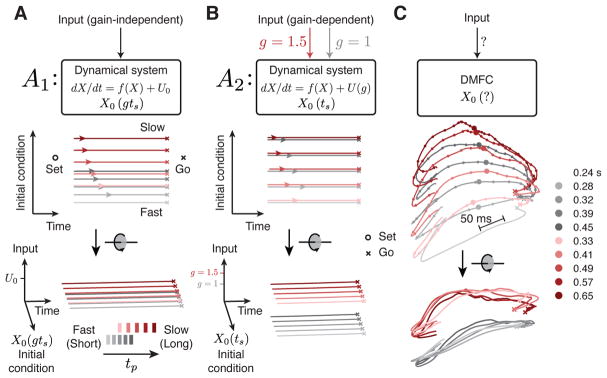
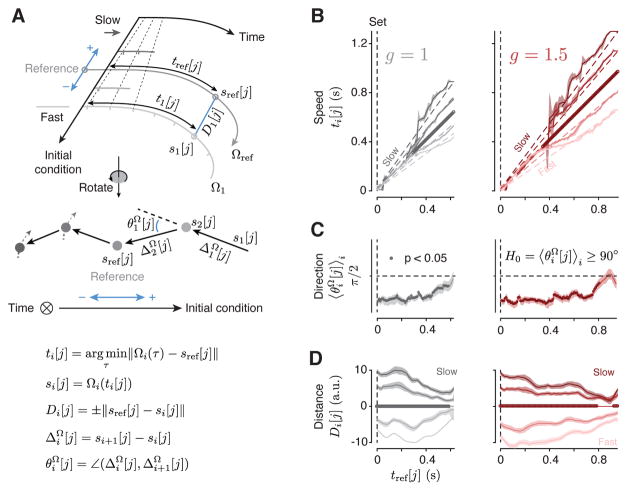
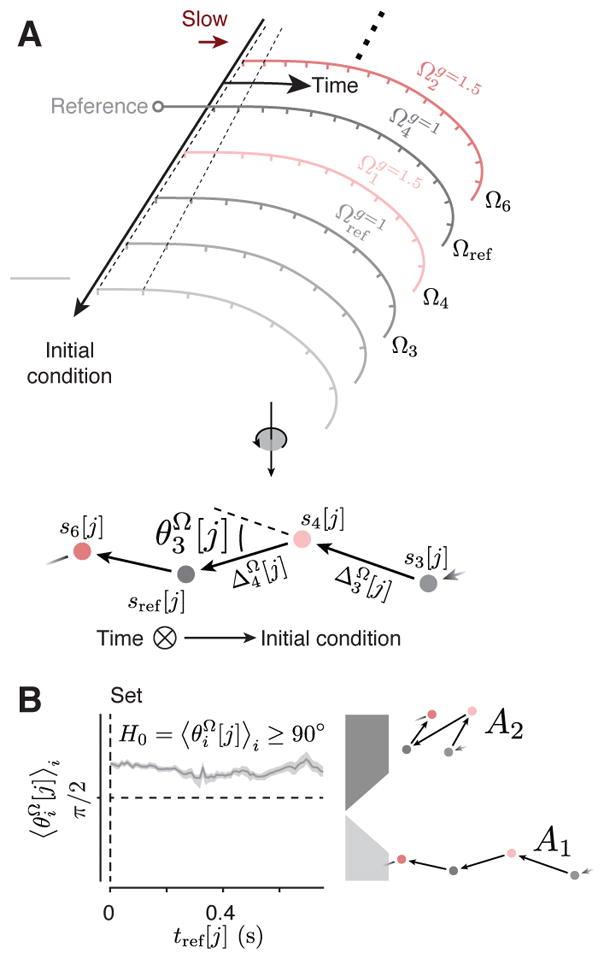
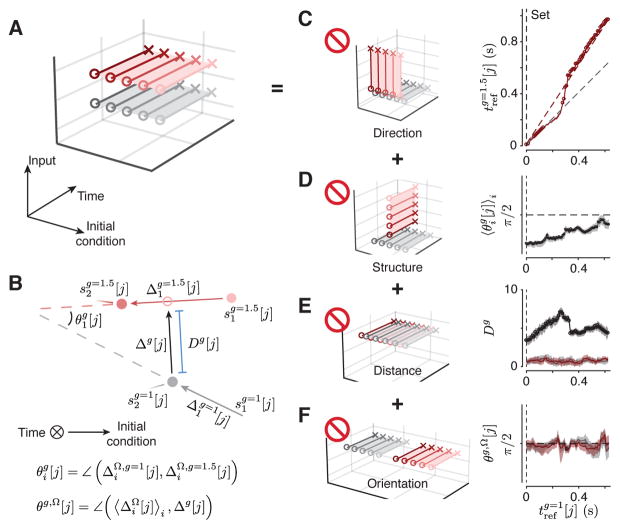
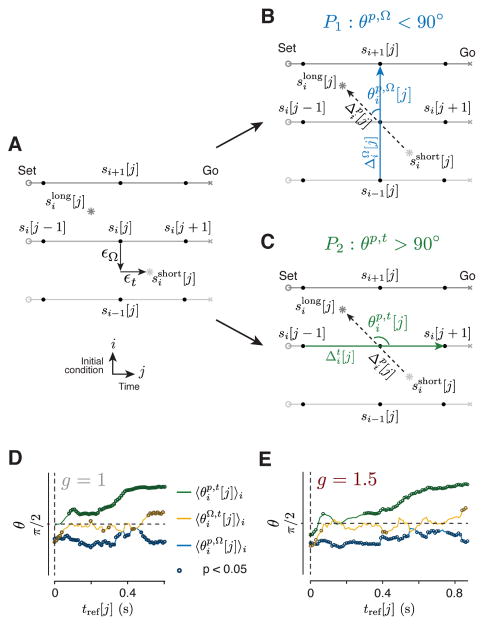
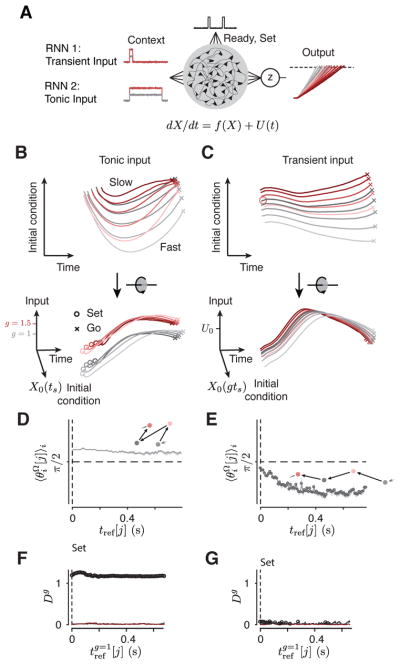
Neural mechanisms that support flexible sensorimotor computations are not well understood. In a dynamical system whose state is determined by interactions among neurons, computations can be rapidly reconfigured by controlling the system's inputs and initial conditions. To investigate whether the brain employs such control mechanisms, we recorded from the dorsomedial frontal cortex of monkeys trained to measure and produce time intervals in two sensorimotor contexts. The geometry of neural trajectories during the production epoch was consistent with a mechanism wherein the measured interval and sensorimotor context exerted control over cortical dynamics by adjusting the system's initial condition and input, respectively. These adjustments, in turn, set the speed at which activity evolved in the production epoch, allowing the animal to flexibly produce different time intervals. These results provide evidence that the language of dynamical systems can be used to parsimoniously link brain activity to sensorimotor computations.
Keywords: Dynamical Systems; cognitive flexibility; electrophysiology; frontal cortex; motor planning; population coding; recurrent neural networks; sensorimotor coordination; timing.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Computation through Cortical Dynamics.Neuron. 2018 Jun 6;98(5):873-875. doi: 10.1016/j.neuron.2018.05.029. Neuron. 2018. PMID: 29879388
Similar articles
-
Bayesian Computation through Cortical Latent Dynamics.Neuron. 2019 Sep 4;103(5):934-947.e5. doi: 10.1016/j.neuron.2019.06.012. Epub 2019 Jul 15. Neuron. 2019. PMID: 31320220 Free PMC article.
-
Neural interactions in the frontal cortex of a behaving monkey: signs of dependence on stimulus context and behavioral state.J Hirnforsch. 1991;32(6):735-43. J Hirnforsch. 1991. PMID: 1821420
-
Neural geometry from mixed sensorimotor selectivity for predictive sensorimotor control.Elife. 2025 May 1;13:RP100064. doi: 10.7554/eLife.100064. Elife. 2025. PMID: 40310450 Free PMC article.
-
Neural Encoding and Representation of Time for Sensorimotor Control and Learning.J Neurosci. 2021 Feb 3;41(5):866-872. doi: 10.1523/JNEUROSCI.1652-20.2020. Epub 2020 Dec 30. J Neurosci. 2021. PMID: 33380468 Free PMC article. Review.
-
Cognition and the single neuron: How cell types construct the dynamic computations of frontal cortex.Curr Opin Neurobiol. 2022 Dec;77:102630. doi: 10.1016/j.conb.2022.102630. Epub 2022 Oct 7. Curr Opin Neurobiol. 2022. PMID: 36209695 Free PMC article. Review.
Cited by
-
Minimally dependent activity subspaces for working memory and motor preparation in the lateral prefrontal cortex.Elife. 2020 Sep 9;9:e58154. doi: 10.7554/eLife.58154. Elife. 2020. PMID: 32902383 Free PMC article.
-
A neural circuit model for human sensorimotor timing.Nat Commun. 2020 Aug 7;11(1):3933. doi: 10.1038/s41467-020-16999-8. Nat Commun. 2020. PMID: 32770038 Free PMC article.
-
Dopamine precursor depletion affects performance and confidence judgements when events are timed from an explicit, but not an implicit onset.Sci Rep. 2023 Dec 11;13(1):21933. doi: 10.1038/s41598-023-47843-w. Sci Rep. 2023. PMID: 38081860 Free PMC article.
-
An abstract categorical decision code in dorsal premotor cortex.Proc Natl Acad Sci U S A. 2022 Dec 13;119(50):e2214562119. doi: 10.1073/pnas.2214562119. Epub 2022 Dec 5. Proc Natl Acad Sci U S A. 2022. PMID: 36469775 Free PMC article.
-
A Neural Population Mechanism for Rapid Learning.Neuron. 2018 Nov 21;100(4):964-976.e7. doi: 10.1016/j.neuron.2018.09.030. Epub 2018 Oct 18. Neuron. 2018. PMID: 30344047 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
