

Lmo2 expression defines tumor cell identity during T-cell leukemogenesis
- PMID: 29880602
- PMCID: PMC6043907
- DOI: 10.15252/embj.201798783
Lmo2 expression defines tumor cell identity during T-cell leukemogenesis
Abstract
The impact of LMO2 expression on cell lineage decisions during T-cell leukemogenesis remains largely elusive. Using genetic lineage tracing, we have explored the potential of LMO2 in dictating a T-cell malignant phenotype. We first initiated LMO2 expression in hematopoietic stem/progenitor cells and maintained its expression in all hematopoietic cells. These mice develop exclusively aggressive human-like T-ALL In order to uncover a potential exclusive reprogramming effect of LMO2 in murine hematopoietic stem/progenitor cells, we next showed that transient LMO2 expression is sufficient for oncogenic function and induction of T-ALL The resulting T-ALLs lacked LMO2 and its target-gene expression, and histologically, transcriptionally, and genetically similar to human LMO2-driven T-ALL We next found that during T-ALL development, secondary genomic alterations take place within the thymus. However, the permissiveness for development of T-ALL seems to be associated with wider windows of differentiation than previously appreciated. Restricted Cre-mediated activation of Lmo2 at different stages of B-cell development induces systematically and unexpectedly T-ALL that closely resembled those of their natural counterparts. Together, these results provide a novel paradigm for the generation of tumor T cells through reprogramming in vivo and could be relevant to improve the response of T-ALL to current therapies.
Keywords: cancer initiation; epigenetic priming; mouse models; oncogenes; stem cells.
© 2018 The Authors. Published under the terms of the CC BY 4.0 license.
Figures
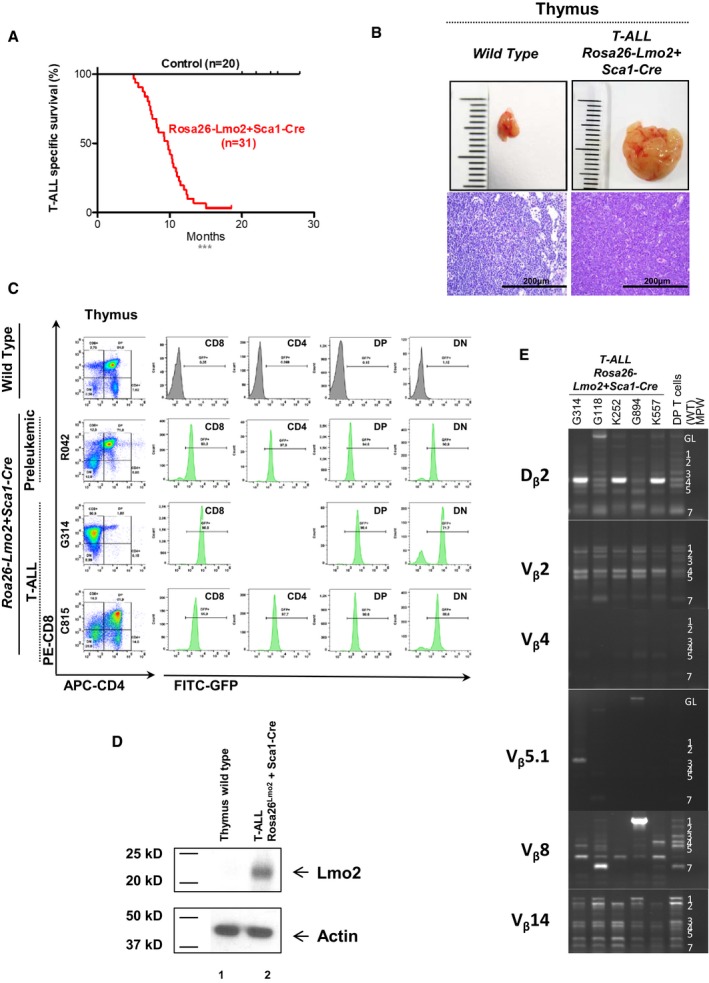
Leukemia‐specific survival of Rosa26‐Lmo2 + Sca1‐Cre mice (red line, n = 31), showing a significantly (log‐rank ***P < 0.0001) shortened lifespan compared to control littermate WT mice (black line, n = 20) as a result of T‐ALL development.
An example of thymomas observed in the Rosa26‐Lmo2 + Sca1‐Cre mice studied. A thymus from a control littermate WT mouse is shown for reference. Hematoxylin and eosin staining showing infiltration of the thymus in Rosa26‐Lmo2 + Sca1‐Cre leukemic mice. Images are photographed at 400× magnification (scale bars: 200 μm).
GFP expression in the pre‐leukemic and leukemic cells from Rosa26‐Lmo2 + Sca1‐Cre mice, respectively. A control littermate WT mouse is shown for reference.
Western blot analysis for Lmo2 and actin in T cells from the thymus of a wild‐type mouse (1) and from the thymus of a Rosa26‐Lmo2 + Sca1‐Cre leukemic mouse (2). Tumoral cells of Rosa26‐Lmo2 + Sca1‐Cre T‐ALL showed expression of the Lmo2 protein.
TCR clonality in Rosa26‐Lmo2 + Sca1‐Cre mice. PCR analysis of TCR gene rearrangements in infiltrated thymuses of diseased Rosa26‐Lmo2 + Sca1‐Cre leukemic mice. Sorted DP T cells from the thymus of healthy mice served as a control for polyclonal TCR rearrangements. Leukemic thymus shows an increased clonality within their TCR repertoire (indicated by the code number of each Rosa26‐Lmo2 + Sca1‐Cre mouse analyzed).
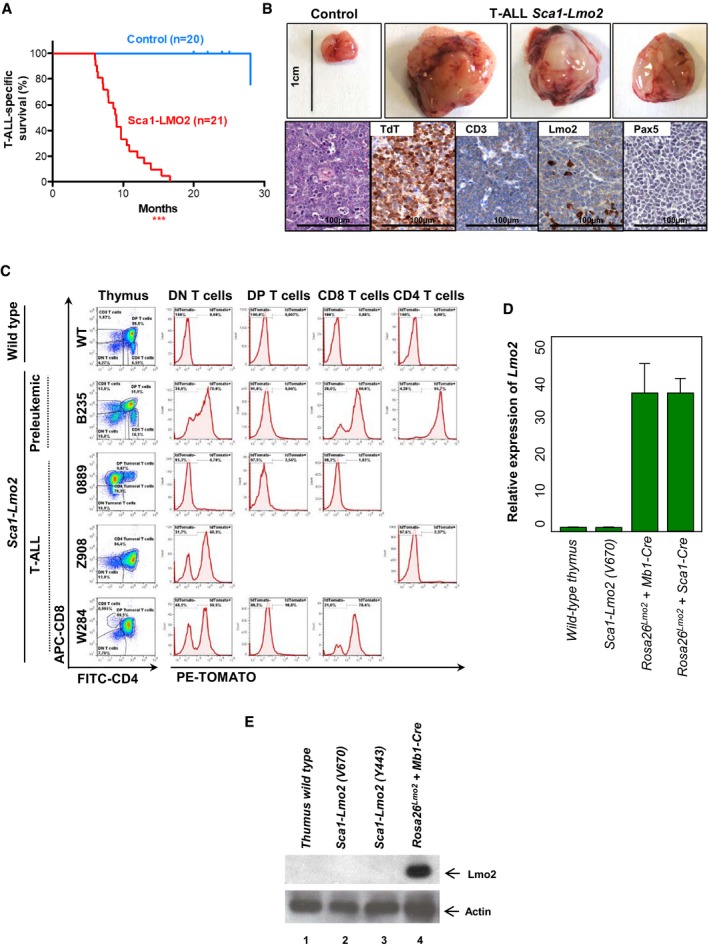
T‐ALL‐specific survival of Sca1‐Lmo2 mice (red line, n = 21), showing a significantly (log‐rank ***P < 0.0001) shortened lifespan compared to control littermate WT mice (blue line, n = 20) as a result of mature T‐cell malignancies.
An example of thymomas observed in 100% (21/21) of the Sca1‐Lmo2 mice studied. A thymus from a control littermate WT mouse is shown for reference. Hematoxylin and eosin staining of tumor‐bearing thymuses from Sca1‐Lmo2 mice shows infiltrate of medium‐sized, relatively uniform lymphoid cells that have a high nuclear/cytoplasmic ratio and immature chromatin with a starry‐sky appearance. Immunohistochemistry shows that tumor T cells from Sca1‐Lmo2 thymomas are defined by the presence of TdT (a marker of T‐cell identity) and CD3 (a marker of immature lymphoid cells), and the absence of Pax5 and Lmo2. Images are representative of ≥ 3 replicates. Images are photographed at 300× magnifications (scale bars: 100 μm).
Flow cytometric analysis of T‐cell subsets in the thymuses of diseased Sca1‐Lmo2 mice. Representative plots of cell subsets from the thymuses are shown. These exhibited the accumulation of DP, CD8, or CD4 single‐positive tumoral T cells. Thymuses from a control littermate WT mouse and a pre‐leukemic Sca1‐Lmo2 mouse are shown for reference. Flow cytometric images are representative of 17 mice analyzed. Tracking of the TdTomato marker for Lmo2 transgene expression in the thymomas of Sca1‐Lmo2 mice shows that tumor T cells are TdTomato‐negative in 100% (17/17) of the Sca1‐Lmo2 mice studied. However, a mosaic of Lmo2 expression remains present within non‐tumoral T‐cell populations (not denoted as tumoral populations). Three plots of cell subsets from the thymuses of diseased Sca1‐Lmo2 mice are shown and are representative of the analysis of 17 diseased Sca1‐Lmo2 mice. TdTomato expression in the thymuses of a control littermate WT mouse and a pre‐leukemic Sca1‐Lmo2 mouse is shown for reference.
Relative expression of Lmo2 in sorted‐purified leukemic Sca1‐Lmo2 cells compared to control thymus wild‐type cells. Leukemic cells of both Rosa26‐Lmo2 + Mb1‐Cre and Rosa26‐Lmo2 + Sca1‐Cre mice were used as a positive control. The fold change in each group, calculated as 2 sample, was compared. Bars represent the mean and the standard deviation of three replicates.
Western blot analysis for Lmo2 and actin in T cells from a wild‐type thymus (1), in sorted‐purified leukemic Sca1‐Lmo2 cells (2, 3) and from the thymus of a Rosa26‐Lmo2 + Mb1‐Cre leukemic mouse (4).
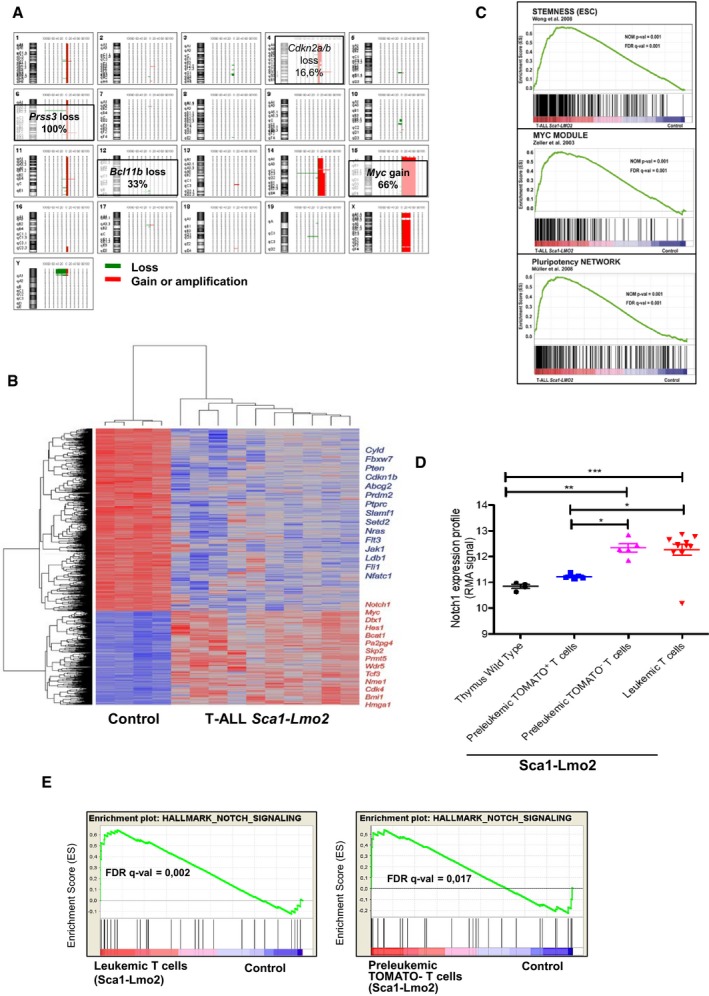
Overview of chromosomal imbalances mapped by 4x180k oligonucleotide aCGH in 12 T‐ALL cases in Sca1‐Lmo2 mice. The 20 chromosome ideograms of T‐ALL Sca1‐Lmo2 mice are shown with DNA deletions drawn as green lines and amplifications or gains as red lines. Selected chromosomal alterations are highlighted.
Genes significantly induced or repressed within tumor T cells of Sca1‐Lmo2 mice in comparison with WT littermates, as determined by significance analysis of microarrays using FDR 1%. Each row represents a separate gene, and each column denotes a separate mRNA sample. The level of expression of each gene in each sample is represented using a red–blue color scale (upregulated genes are displayed in red and downregulated genes in blue). Selected genes are highlighted.
GSEA of the transcriptional signatures within tumor T cells compared with control WT littermates. Gene expression data from Sca1‐Lmo2 tumor T cells showed significant enrichment in embryonic stem cell genes (Wong et al, 2008) (GSEA FDR q‐value = 0.001), Myc target genes (Zeller et al, 2003) (GSEA FDR q‐value = 0.001), and pluripotency genes (Muller et al, 2008) (GSEA FDR q‐value=0.001).
Notch1 expression profile in control WT T cells, pre‐leukemic tomato+ T cells, pre‐leukemic tomato− T cells, and leukemic T cells. The statistical test used was Mann–Whitney U‐test: wild‐type thymus versus leukemic T cells (***P < 0.001), wild‐type thymus versus pre‐leukemic tomato− T cells (**P = 0.0031), pre‐leukemic tomato+ T cells versus leukemic T cells (*P = 0.0127), and pre‐leukemic tomato+ T cells versus pre‐leukemic tomato− T cells (*P = 0.0159). Error bars represent the mean ± SEM.
GSEA of the Notch signaling in leukemic and tomato− cells (GSEA FDR q‐value = 0.002, 0.017, respectively).
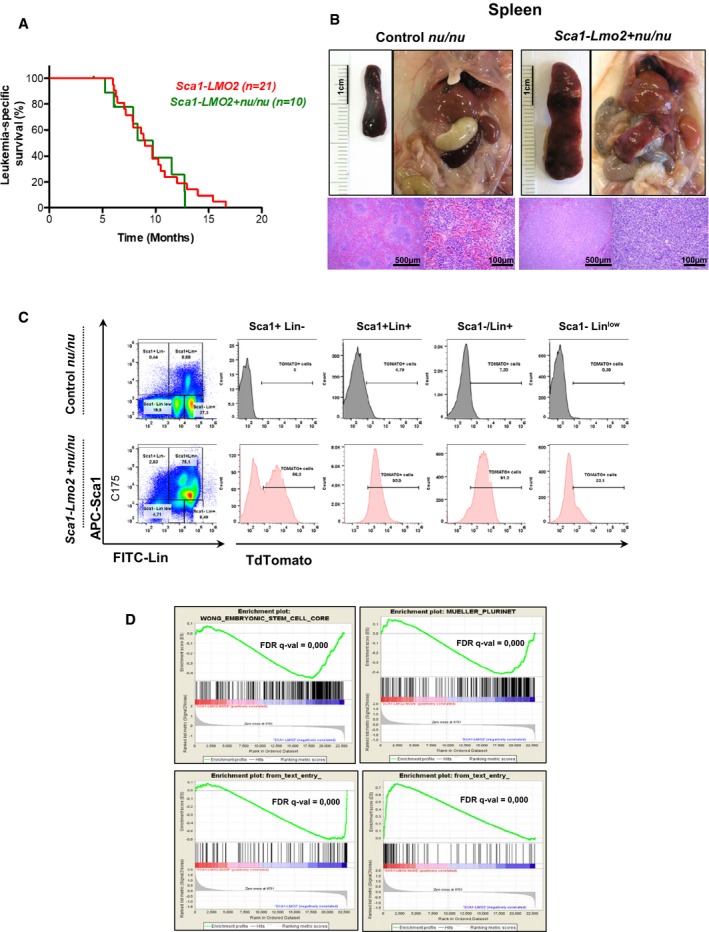
Leukemia‐specific survival of Sca1‐Lmo2 + nu/nu mice (green line, n = 10), showing a similar shortened lifespan compared to Sca1‐Lmo2 mice (red line, n = 21) as a result of leukemia development.
An example of splenomegaly observed in Sca1‐Lmo2 + nu/nu mice studied pointing out by an arrowhead. Hematoxylin and eosin staining showing infiltration of spleen from Sca1‐Lmo2 + nu/nu leukemic mice. A spleen from a control littermate nu/nu mouse is shown for reference.
TdTomato expression in the leukemic cells from Sca1‐Lmo2 + nu/nu mouse. A control littermate nu/nu mouse is shown for reference.
GSEA of the transcriptional signatures within tumor cells of Sca1‐Lmo2 + nu/nu mice compared to tumor T cells of Sca1‐Lmo2 mice. Gene expression data from Sca1‐Lmo2 + nu/nu tumor cells showed significant enrichment of embryonic stem cell genes (GSEA FDR q‐value = 0.000), pluripotency genes (GSEA FDR q‐value = 0.000), genes upregulated in human ETP T‐ALL (GSEA FDR q‐value = 0.000), and genes downregulated in human ETP T‐ALL (GSEA FDR q‐value = 0.000).
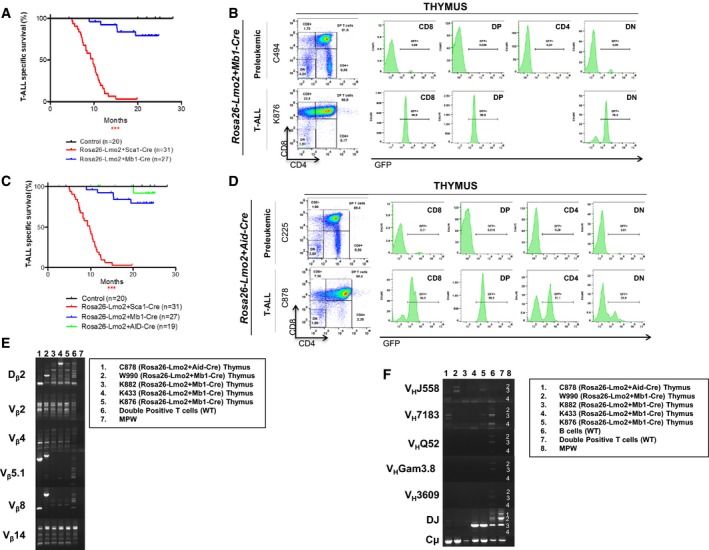
Leukemia‐specific survival of Rosa26‐Lmo2 + Mb1‐Cre mice (blue line, n = 27), showing a significantly (log‐rank ***P < 0.0328) shortened lifespan compared to control littermate WT mice (black line, n = 20) as a result of T‐ALL development. The latency of Rosa26‐Lmo2 + Mb1‐Cre T‐ALL is higher than that of Rosa26‐Lmo2 + Sca1‐Cre T‐ALL.
GFP expression in the pre‐leukemic and leukemic cells from Rosa26‐Lmo2 + Mb1‐Cre mice, respectively.
Leukemia‐specific survival of Rosa26‐Lmo2 + Aid‐Cre mice (green line, n = 19), not showing a significantly (log‐rank ***P < 0.3173) shortened lifespan compared to control littermate WT mice (black line, n = 20). The latency of Rosa26‐Lmo2 + Aid‐Cre T‐ALL is higher than that of Rosa26‐Lmo2 + Sca1‐Cre and Rosa26‐Lmo2 + Mb1‐Cre T‐ALLs.
GFP expression in the pre‐leukemic and leukemic cells from Rosa26‐Lmo2 + Aid‐Cre mice, respectively.
TCR clonality in Rosa26‐Lmo2 + Aid‐Cre and Rosa26‐Lmo2 + Mb1‐Cre mice. PCR analysis of TCR gene rearrangements in infiltrated thymuses of diseased Rosa26‐Lmo2 + Aid‐Cre and Rosa26‐Lmo2 + Mb1‐Cre leukemic mice. Sorted DP T cells from the thymus of healthy mice served as a control for polyclonal TCR rearrangements. Leukemic thymus shows an increased clonality within their TCR repertoire (indicated by the code number of each mouse analyzed).
BCR clonality in Rosa26‐Lmo2 + Aid‐Cre and Rosa26‐Lmo2 + Mb1‐Cre mice. PCR analysis of BCR gene rearrangements in infiltrated thymuses of diseased Rosa26‐Lmo2 + Aid‐Cre and Rosa26‐Lmo2 + Mb1‐Cre leukemic mice. Sorted CD19+ B cells (B cells) from spleens of healthy mice serve as a control for polyclonal BCR rearrangements. DP T cells from the thymus of healthy mice served as a negative control. Leukemic thymus shows an increased clonality within their BCR repertoire (indicated by the code number of each mouse analyzed).
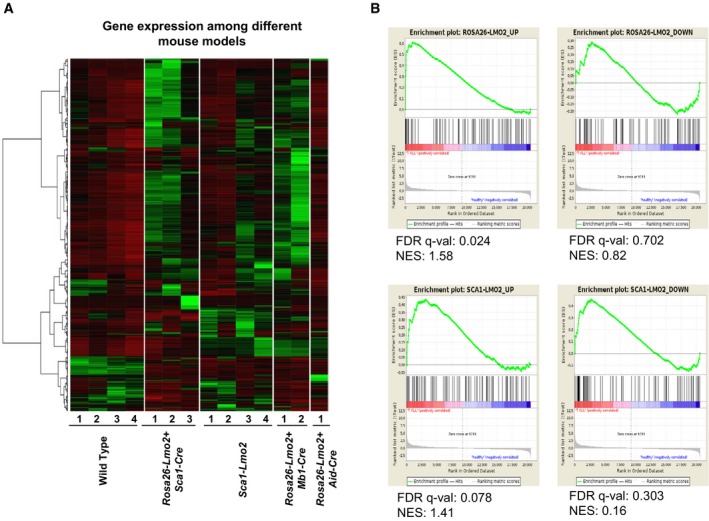
Gene expression of Rosa26‐Lmo2 + Sca1‐Cre, Sca1‐Lmo2, Rosa26‐Lmo2 + Mb1‐Cre, and Rosa26‐Lmo2 + Aid‐Cre with WT thymus as comparison. The 500 genes with the highest variance among the murine groups were chosen, and their corresponding FPKM values transformed to standard scores for visualization. [Row clustering was conducted with the ward.D method.]
Gene set enrichment analysis (GSEA) of Rosa26‐Lmo2 + Sca1‐Cre and Sca1‐Lmo2 mouse‐based gene signatures, against a human T‐ALL childhood expression set with healthy controls. Mouse‐based signatures consist of the 100 most up‐ and downregulated human homologue genes, as identified in a differential gene expression analysis between Rosa26‐Lmo2 + Sca1‐Cre versus WT and Sca1‐Lmo2 versus WT, respectively.
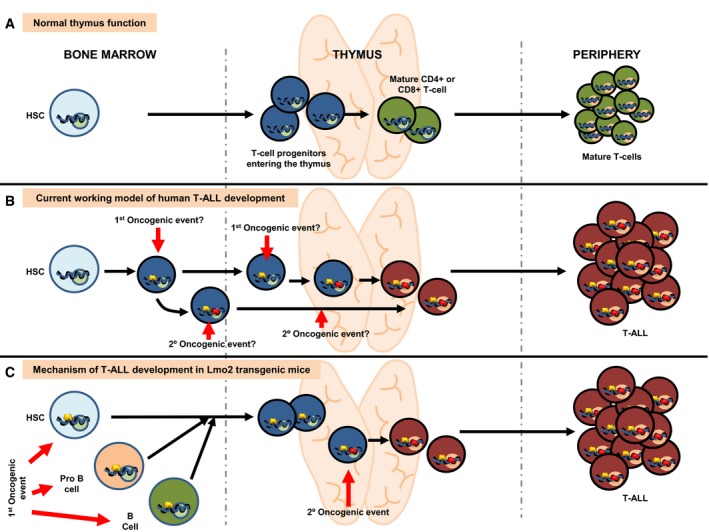
Normal lymphoid development in human and mice. Blue circles represent normal gene regulatory events (activating or repressing) happening during T lymphocyte development. Green circles represent normal gene regulation events happening during terminal differentiation.
Current working model for the development of T‐ALL in humans. The existence of dormant alterations previous to the terminal differentiation is unknown. The nature of both the cancer cell‐of‐origin and the cellular place where the second hit is taking place is therefore unknown.
Mechanism of T‐ALL development in Sca1‐Lmo2 transgenic mice. Open yellow circles represent latent epigenetic regulatory events caused by Sca1‐driven expression of Lmo2. These epigenetic marks do not interfere with normal T‐cell development but become active (either activating or repressing) in the process of terminal differentiation when the second hit appears within the thymus, thus leading to the appearance of tumor T cells. According to this model, tumor T cell is the result of a cell reprogramming process that can be initiated even in committed B cells (see text for details).
References
-
- de Boer J, Yeung J, Ellu J, Ramanujachar R, Bornhauser B, Solarska O, Hubank M, Williams O, Brady HJ (2011) The E2A‐HLF oncogenic fusion protein acts through Lmo2 and Bcl‐2 to immortalize hematopoietic progenitors. Leukemia 25: 321–330 - PubMed
-
- Boiers C, Carrelha J, Lutteropp M, Luc S, Green JC, Azzoni E, Woll PS, Mead AJ, Hultquist A, Swiers G, Perdiguero EG, Macaulay IC, Melchiori L, Luis TC, Kharazi S, Bouriez‐Jones T, Deng Q, Ponten A, Atkinson D, Jensen CT et al (2013) Lymphomyeloid contribution of an immune‐restricted progenitor emerging prior to definitive hematopoietic stem cells. Cell Stem Cell 13: 535–548 - PubMed
-
- Boiers C, Richardson SE, Laycock E, Zriwil A, Turati VA, Brown J, Wray JP, Wang D, James C, Herrero J, Sitnicka E, Karlsson S, Smith AJH, Jacobsen SEW, Enver T (2018) A human IPS model implicates embryonic B‐myeloid fate restriction as developmental susceptibility to B Acute lymphoblastic leukemia‐associated ETV6‐RUNX1. Dev Cell 44: 362–377 e367 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
