

Obesity treatment by epigallocatechin-3-gallate-regulated bile acid signaling and its enriched Akkermansia muciniphila
- PMID: 29882708
- PMCID: PMC6219838
- DOI: 10.1096/fj.201800370R
Obesity treatment by epigallocatechin-3-gallate-regulated bile acid signaling and its enriched Akkermansia muciniphila
Abstract
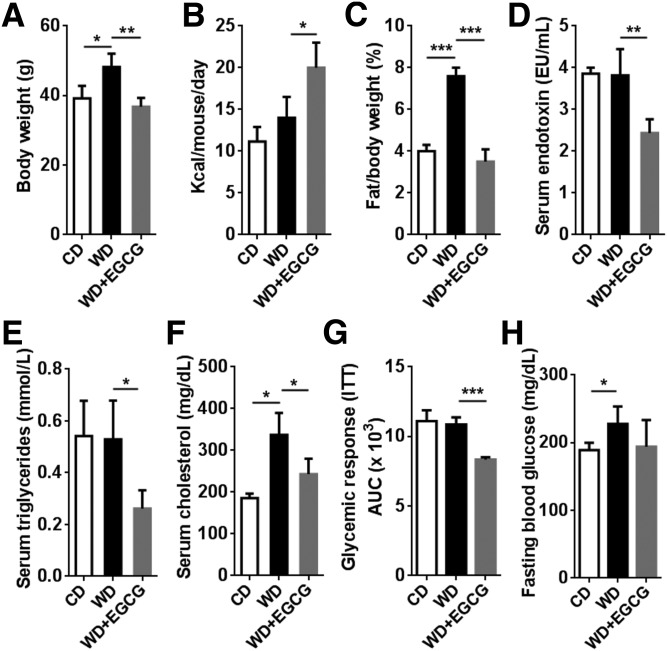
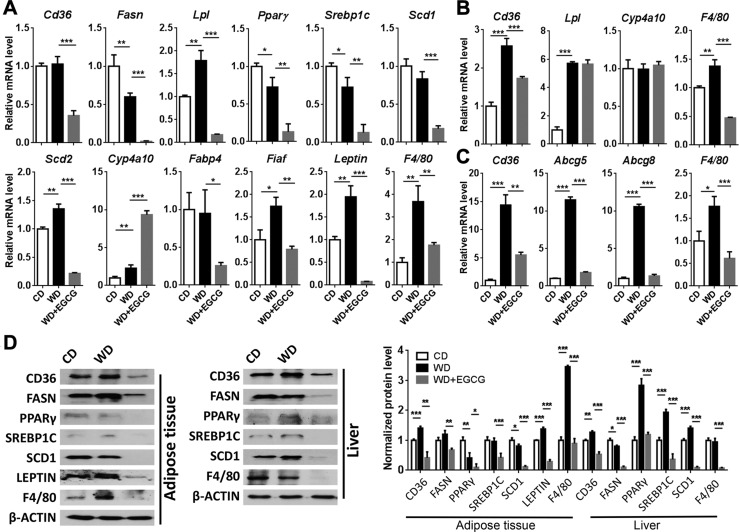
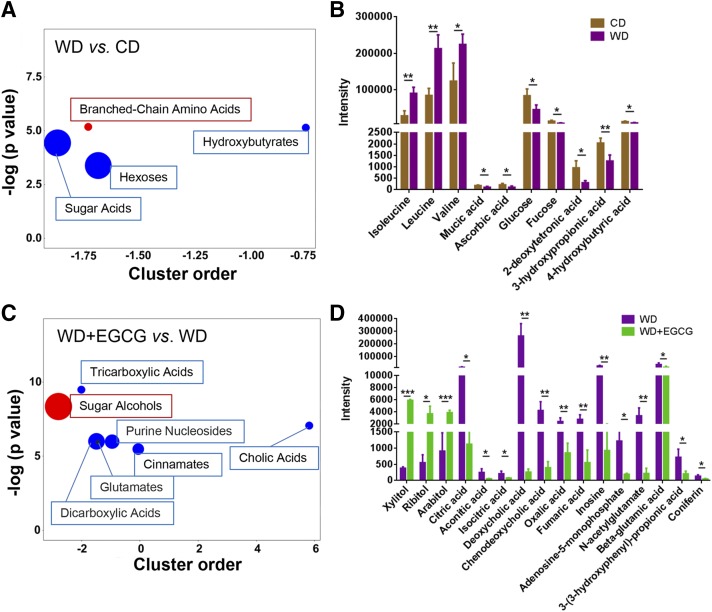
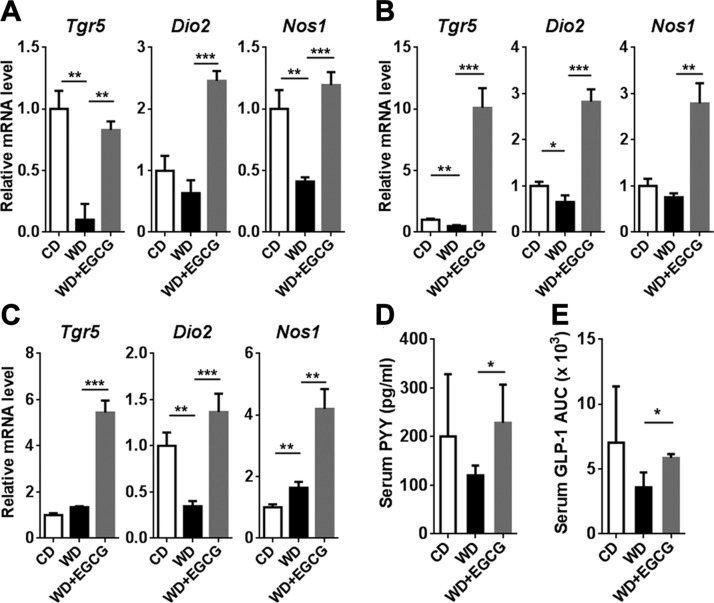
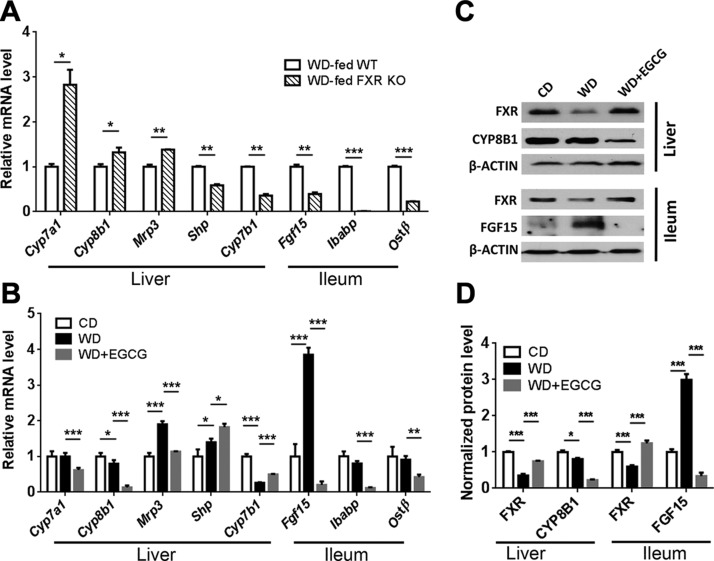
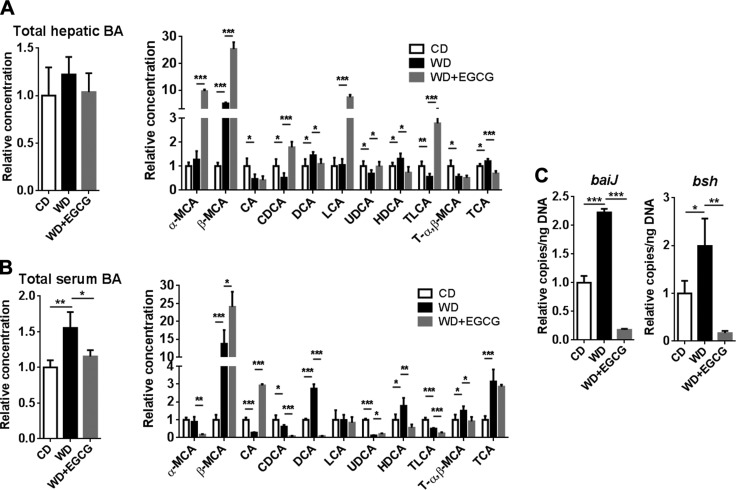
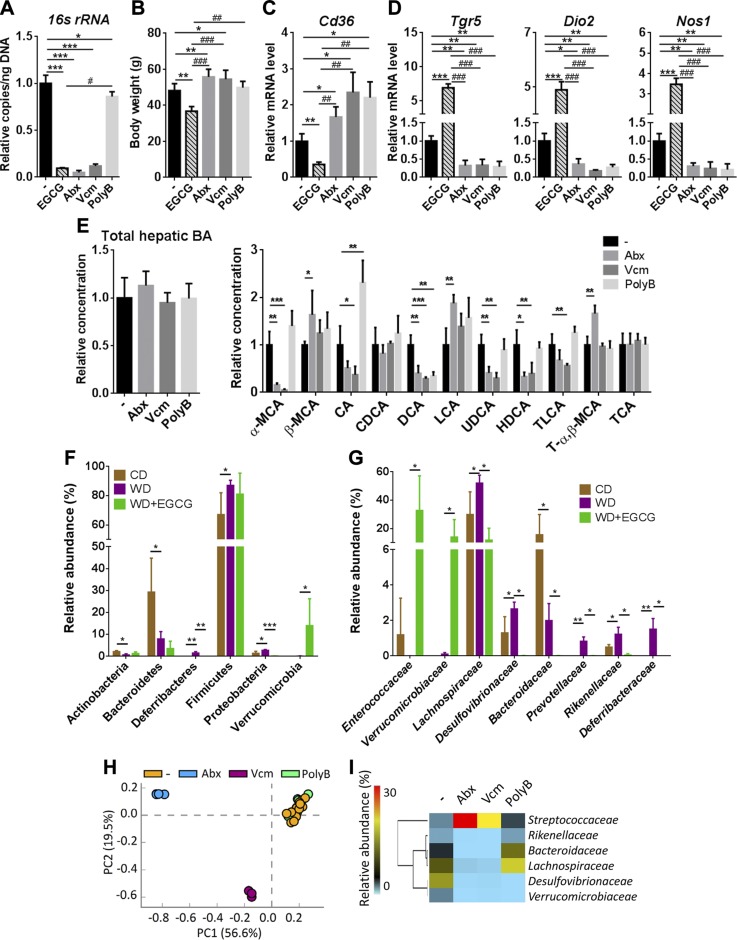
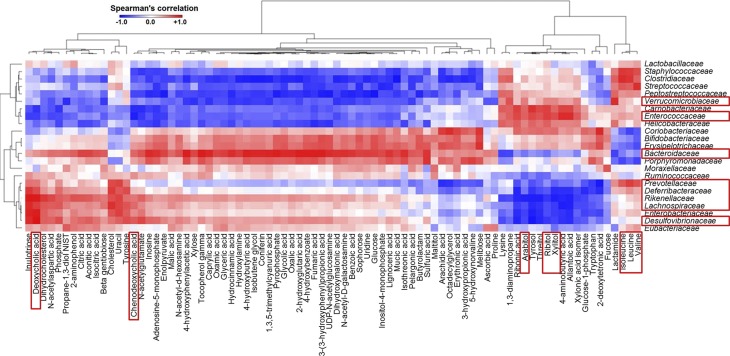
Dysregulated bile acid (BA) synthesis is accompanied by dysbiosis, leading to compromised metabolism. This study analyzes the effect of epigallocatechin-3-gallate (EGCG) on diet-induced obesity through regulation of BA signaling and gut microbiota. The data revealed that EGCG effectively reduced diet-increased obesity, visceral fat, and insulin resistance. Gene profiling data showed that EGCG had a significant impact on regulating genes implicated in fatty acid uptake, adipogenesis, and metabolism in the adipose tissue. In addition, metabolomics analysis revealed that EGCG altered the lipid and sugar metabolic pathways. In the intestine, EGCG reduced the FXR agonist chenodeoxycholic acid, as well as the FXR-regulated pathway, suggesting intestinal FXR deactivation. However, in the liver, EGCG increased the concentration of FXR and TGR-5 agonists and their regulated signaling. Furthermore, our data suggested that EGCG activated Takeda G protein receptor (TGR)-5 based on increased GLP-1 release and elevated serum PYY level. EGCG and antibiotics had distinct antibacterial effects. They also differentially altered body weight and BA composition. EGCG, but not antibiotics, increased Verrucomicrobiaceae, under which EGCG promoted intestinal bloom of Akkermansia muciniphila. Excitingly, A. muciniphila was as effective as EGCG in treating diet-induced obesity. Together, EGCG shifts gut microbiota and regulates BA signaling thereby having a metabolic beneficial effect.-Sheng, L., Jena, P. K., Liu, H.-X., Hu, Y., Nagar, N., Bronner, D. N., Settles, M. L., Bäumler, A. J. Wan, Y.-J. Y. Obesity treatment by epigallocatechin-3-gallate-regulated bile acid signaling and its enriched Akkermansia muciniphila.
Keywords: bile acid receptor; catechin; gut microbiota; probiotics; tea.
Conflict of interest statement
The authors thank Dr. Frank J. Gonzalez [U.S. National Institutes of Health (NIH) National Cancer Institute] for providing FXR KO mice, and Dr. Mel Campbell (University of California, Davis) for editing the manuscript. This study was supported by NIH National Cancer Institute Grants U01CA179582 and R01CA222490 (to Y.-J. Y. Wan). The authors declare no conflicts of interest.
Figures
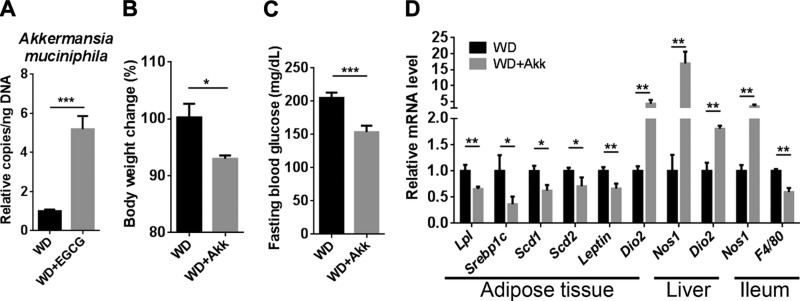
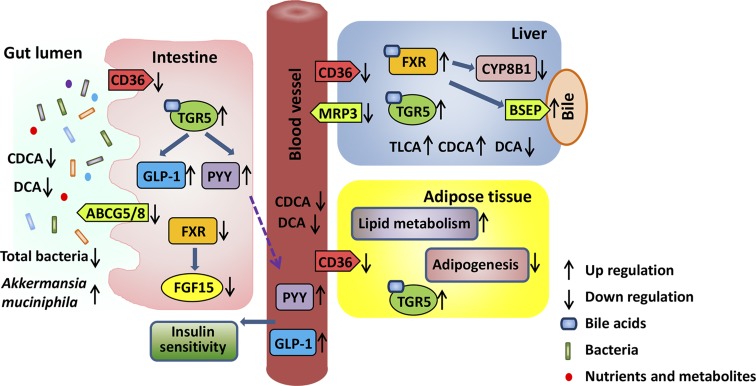
References
-
- Weber J. M., Ruzindana-Umunyana A., Imbeault L., Sircar S. (2003) Inhibition of adenovirus infection and adenain by green tea catechins. Antiviral Res. 58, 167–173 - PubMed
-
- Weinreb O., Mandel S., Amit T., Youdim M. B. (2004) Neurological mechanisms of green tea polyphenols in Alzheimer’s and Parkinson’s diseases. J. Nutr. Biochem. 15, 506–516 - PubMed
-
- Raederstorff D. G., Schlachter M. F., Elste V., Weber P. (2003) Effect of EGCG on lipid absorption and plasma lipid levels in rats. J. Nutr. Biochem. 14, 326–332 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
