

Parallel Processing of Rod and Cone Signals: Retinal Function and Human Perception
- PMID: 29883274
- PMCID: PMC6153147
- DOI: 10.1146/annurev-vision-091517-034055
Parallel Processing of Rod and Cone Signals: Retinal Function and Human Perception
Abstract
We know a good deal about the operation of the retina when either rod or cone photoreceptors provide the dominant input (i.e., under very dim or very bright conditions). However, we know much less about how the retina operates when rods and cones are coactive (i.e., under intermediate lighting conditions, such as dusk). Such mesopic conditions span 20-30% of the light levels over which vision operates and encompass many situations in which vision is essential (e.g., driving at night). These lighting conditions are challenging because rod and cone signals differ substantially: Rod responses are nearing saturation, while cone responses are weak and noisy. A rich history of perceptual studies guides our investigation of how the retina operates under mesopic conditions and in doing so provides a powerful opportunity to link general issues about parallel processing in neural circuits with computation and perception. We review some of the successes and challenges in understanding the retinal basis of perceptual rod-cone interactions.
Keywords: circuit; cone; parallel processing; perception; retina; rod.
Figures
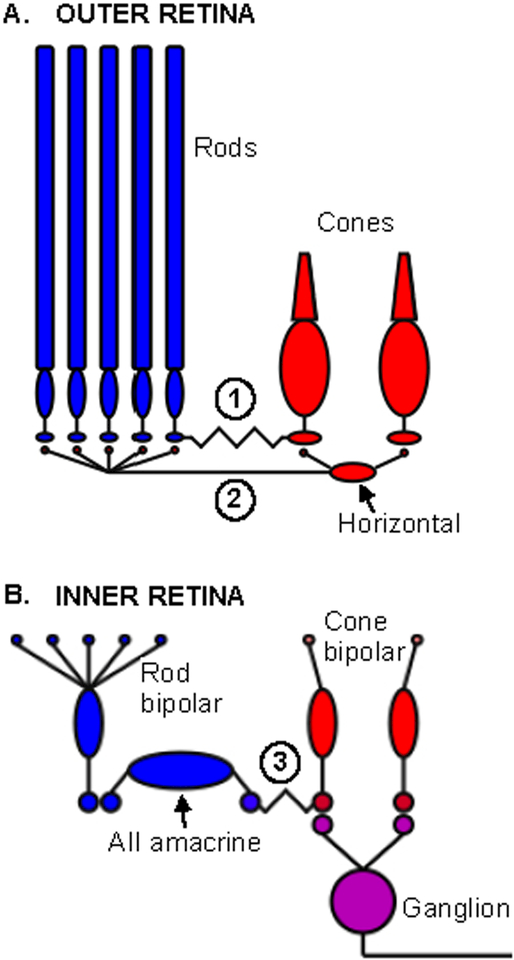
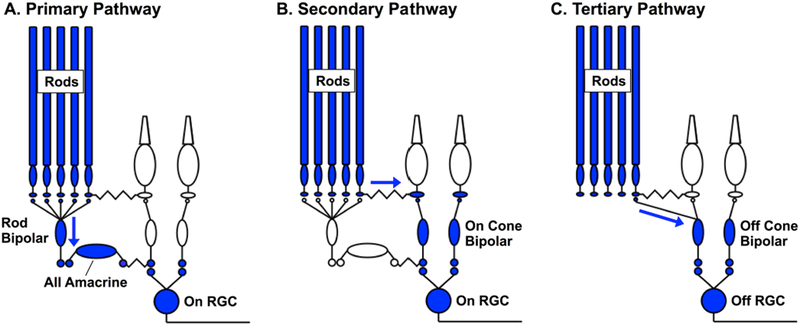
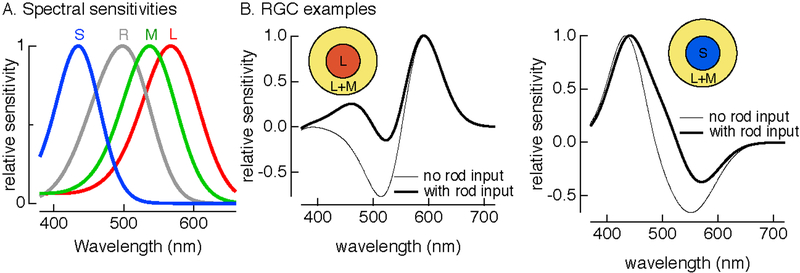
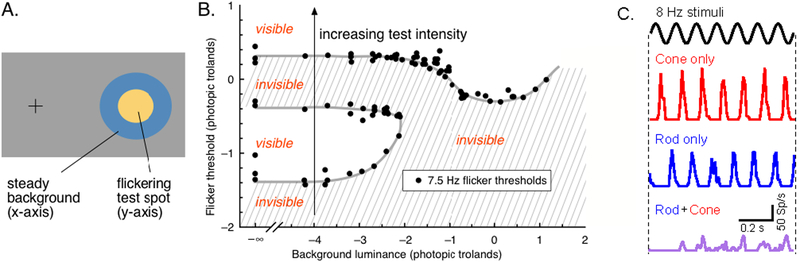
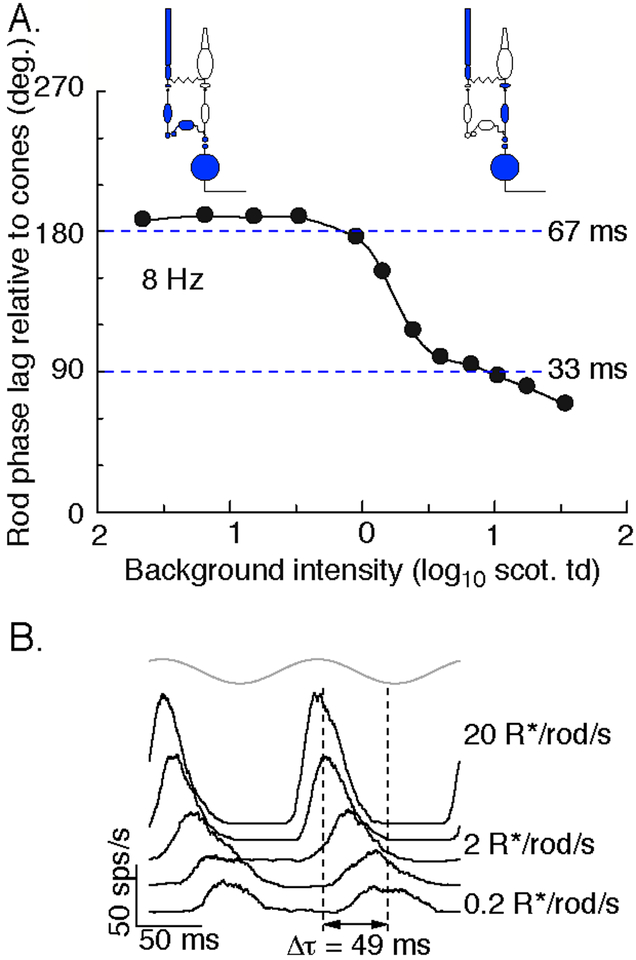
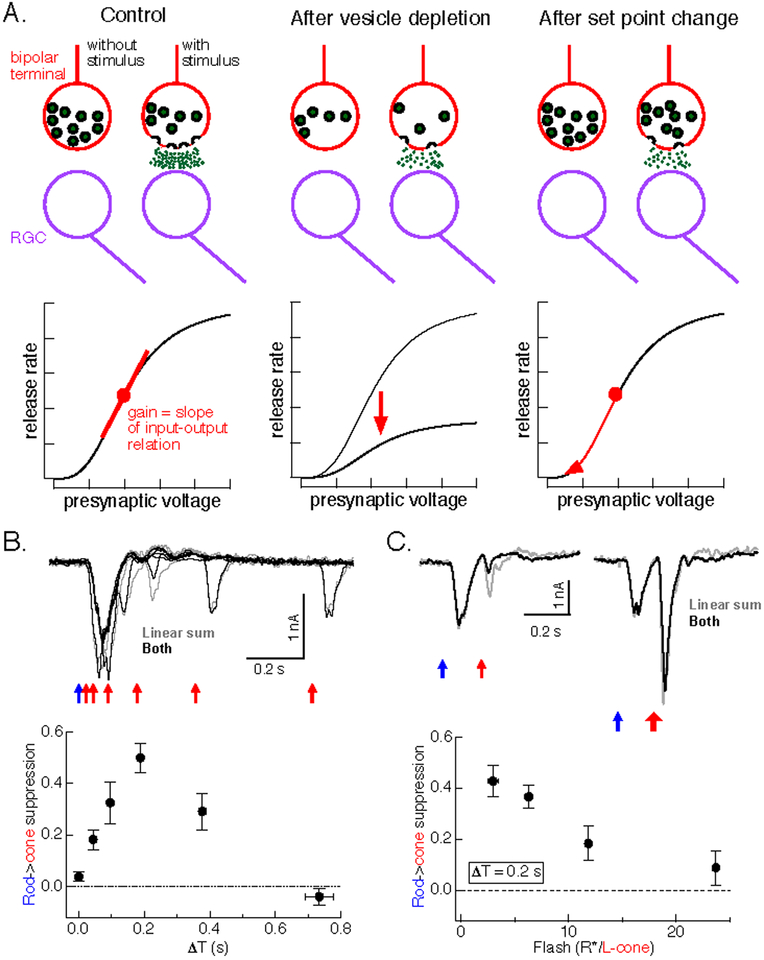
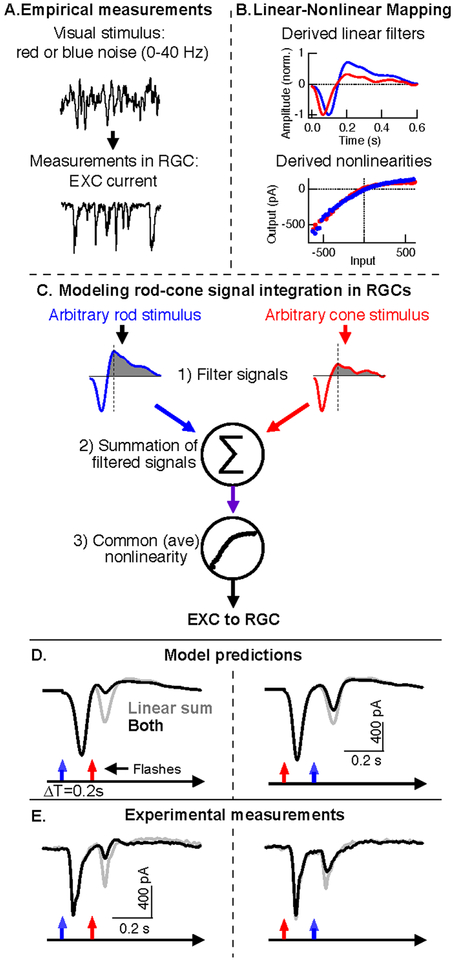
References
-
- Barlow HB, Levick WR & Yoon M (1971) Responses to single quanta of light in retinal ganglion cells of the cat. Vision Res, Suppl 3, 87–101. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
