

Sequence, Structure, and Context Preferences of Human RNA Binding Proteins
- PMID: 29883606
- PMCID: PMC6062212
- DOI: 10.1016/j.molcel.2018.05.001
Sequence, Structure, and Context Preferences of Human RNA Binding Proteins
Abstract
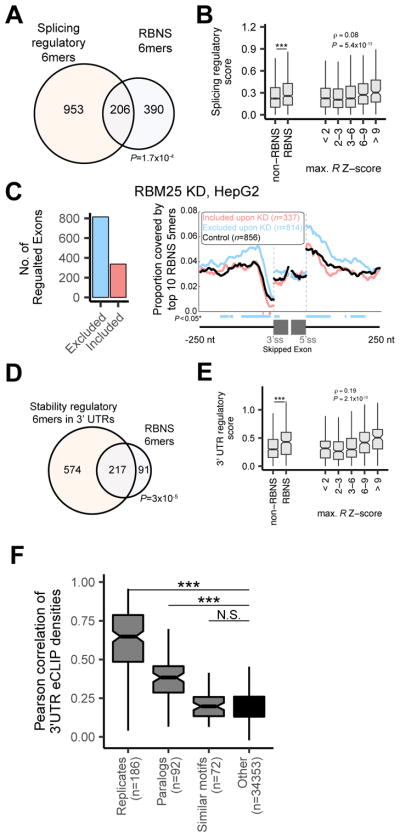
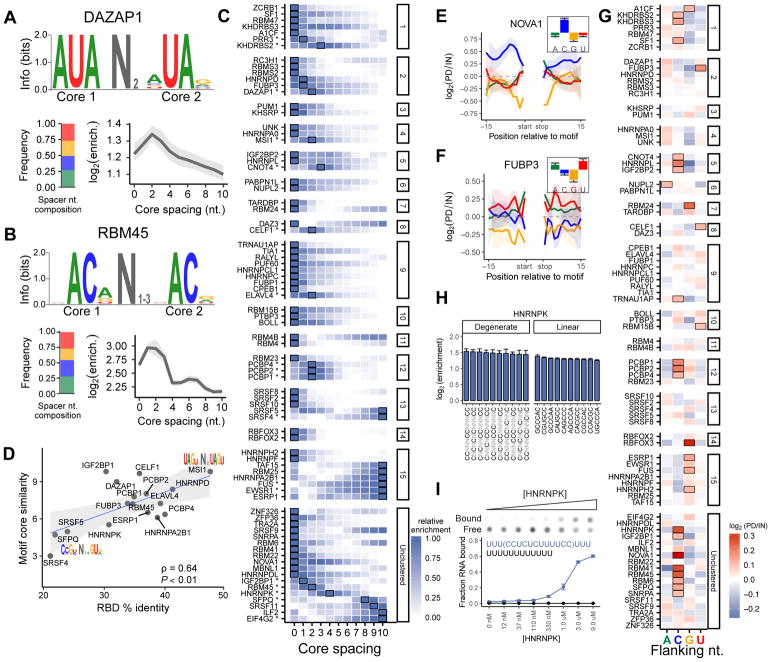
RNA binding proteins (RBPs) orchestrate the production, processing, and function of mRNAs. Here, we present the affinity landscapes of 78 human RBPs using an unbiased assay that determines the sequence, structure, and context preferences of these proteins in vitro by deep sequencing of bound RNAs. These data enable construction of "RNA maps" of RBP activity without requiring crosslinking-based assays. We found an unexpectedly low diversity of RNA motifs, implying frequent convergence of binding specificity toward a relatively small set of RNA motifs, many with low compositional complexity. Offsetting this trend, however, we observed extensive preferences for contextual features distinct from short linear RNA motifs, including spaced "bipartite" motifs, biased flanking nucleotide composition, and bias away from or toward RNA structure. Our results emphasize the importance of contextual features in RNA recognition, which likely enable targeting of distinct subsets of transcripts by different RBPs that recognize the same linear motif.
Keywords: KH domain; Pum domain; RBNS; RNA binding protein; RNA context; RNA recognition motif; RNA secondary structure; alternative splicing; mRNA stability; zinc finger.
Copyright © 2018 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Botti V, McNicoll F, Steiner MC, Richter FM, Solovyeva A, Wegener M, Schwich OD, Poser I, Zarnack K, Wittig I, Neugebauer KM, Müller-McNicoll M. Cellular differentiation state modulates the mRNA export activity of SR proteins. J Cell Biol. 2017;216:1993–2009. doi: 10.1083/jcb.201610051. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
