

Higher-Order Inter-chromosomal Hubs Shape 3D Genome Organization in the Nucleus
- PMID: 29887377
- PMCID: PMC6548320
- DOI: 10.1016/j.cell.2018.05.024
Higher-Order Inter-chromosomal Hubs Shape 3D Genome Organization in the Nucleus
Abstract
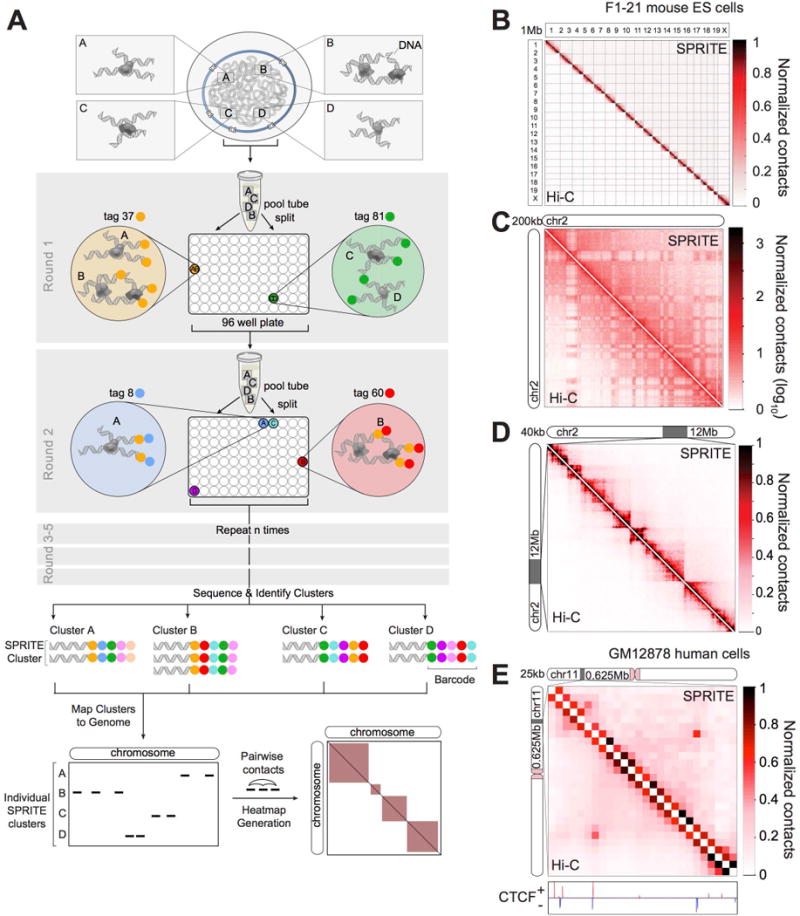
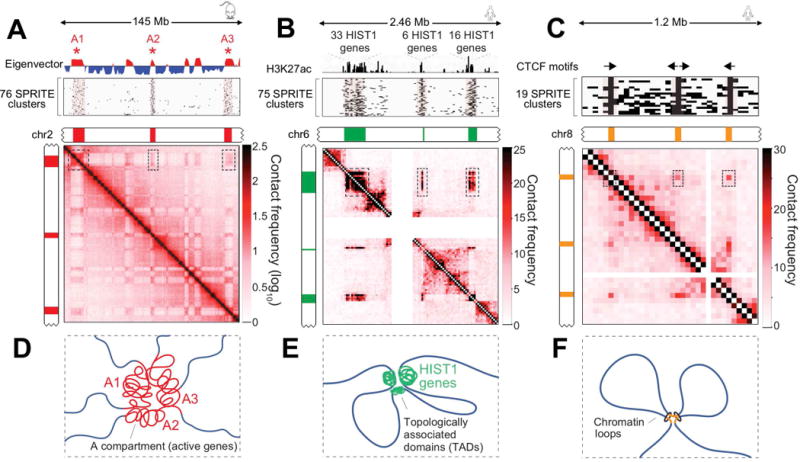
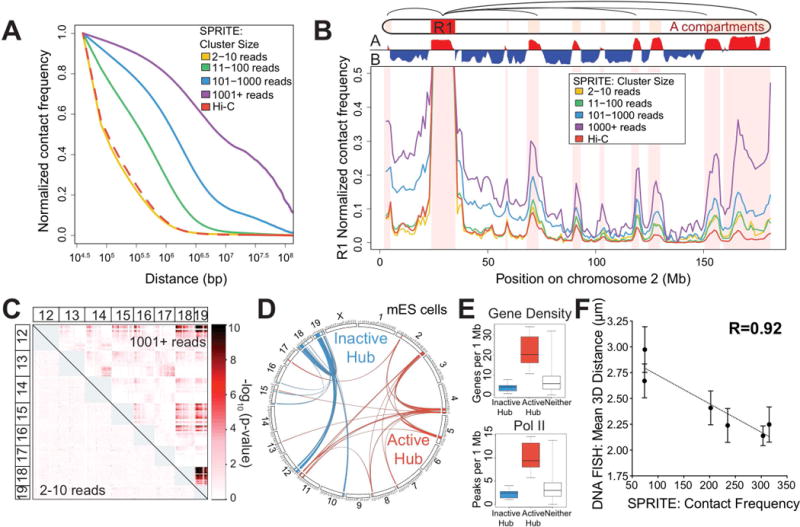
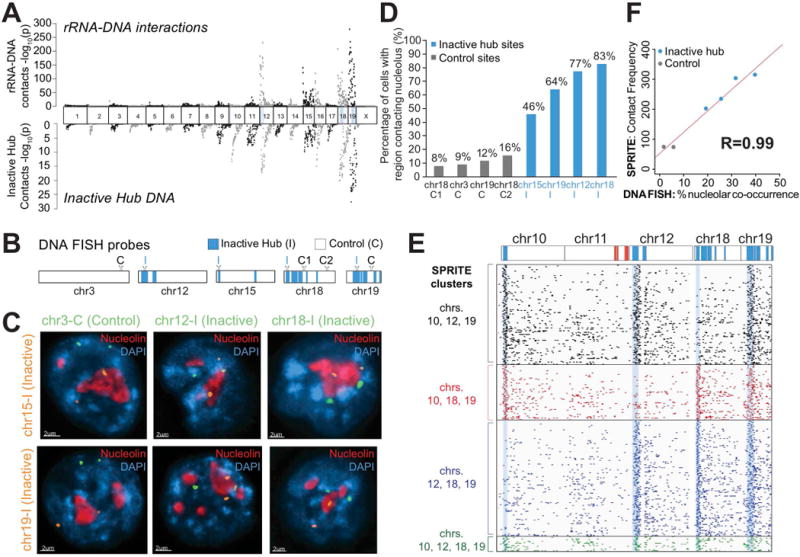
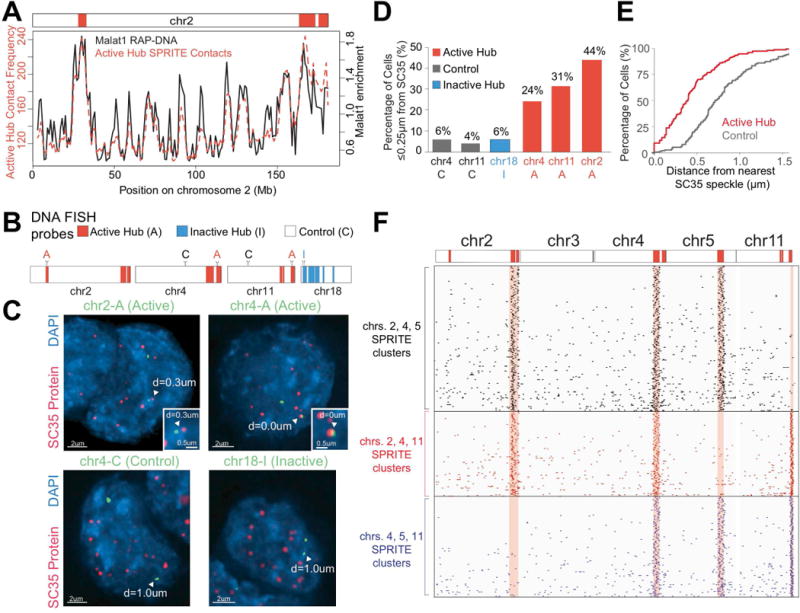
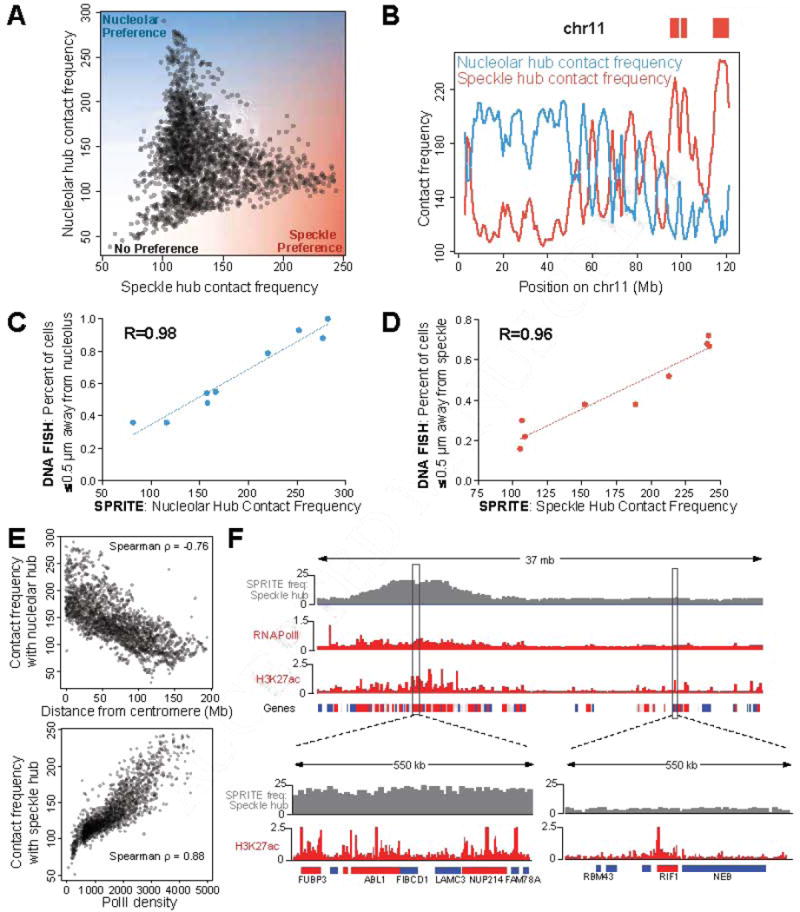
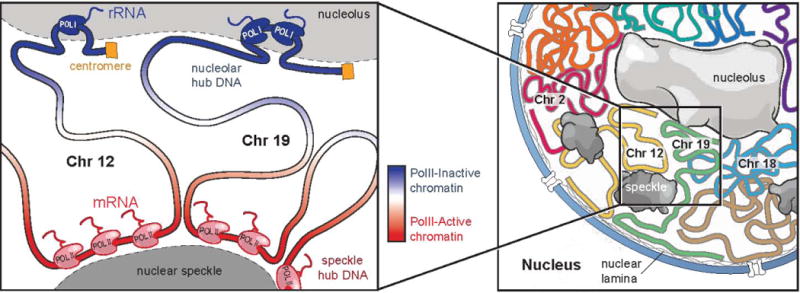
Eukaryotic genomes are packaged into a 3-dimensional structure in the nucleus. Current methods for studying genome-wide structure are based on proximity ligation. However, this approach can fail to detect known structures, such as interactions with nuclear bodies, because these DNA regions can be too far apart to directly ligate. Accordingly, our overall understanding of genome organization remains incomplete. Here, we develop split-pool recognition of interactions by tag extension (SPRITE), a method that enables genome-wide detection of higher-order interactions within the nucleus. Using SPRITE, we recapitulate known structures identified by proximity ligation and identify additional interactions occurring across larger distances, including two hubs of inter-chromosomal interactions that are arranged around the nucleolus and nuclear speckles. We show that a substantial fraction of the genome exhibits preferential organization relative to these nuclear bodies. Our results generate a global model whereby nuclear bodies act as inter-chromosomal hubs that shape the overall packaging of DNA in the nucleus.
Keywords: Nuclear structure; RNA DNA interactions; SPRITE; genome structure; higher-order nuclear structure; multi-way interactions; nuclear organization; nuclear speckle; nucleolus.
Copyright © 2018 Elsevier Inc. All rights reserved.
Conflict of interest statement
S.Q. and M.G. are inventors on a provisional patent on the SPRITE method.
Figures
Comment in
-
Chromosome interaction hubs around nuclear bodies.Nat Rev Genet. 2018 Aug;19(8):470-471. doi: 10.1038/s41576-018-0026-x. Nat Rev Genet. 2018. PMID: 29884879 No abstract available.
-
3D interaction hubs.Nat Chem Biol. 2018 Aug;14(8):745. doi: 10.1038/s41589-018-0112-6. Nat Chem Biol. 2018. PMID: 30018419 No abstract available.
-
SPRITE maps the 3D genome.Nat Methods. 2018 Aug;15(8):572. doi: 10.1038/s41592-018-0092-1. Nat Methods. 2018. PMID: 30065384 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
