

Human CD30+ B cells represent a unique subset related to Hodgkin lymphoma cells
- PMID: 29889102
- PMCID: PMC6025985
- DOI: 10.1172/JCI95993
Human CD30+ B cells represent a unique subset related to Hodgkin lymphoma cells
Abstract
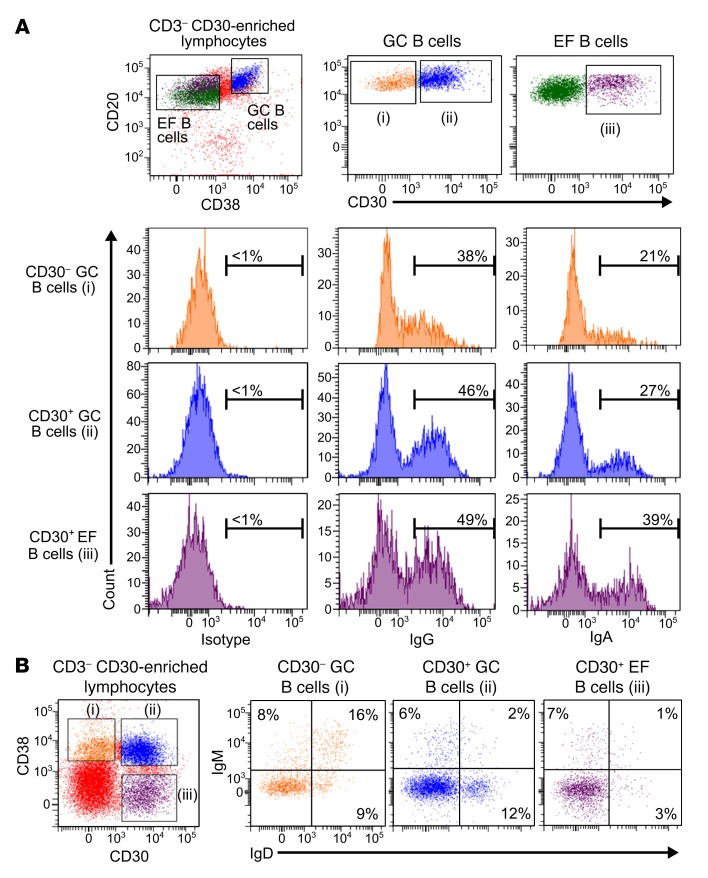
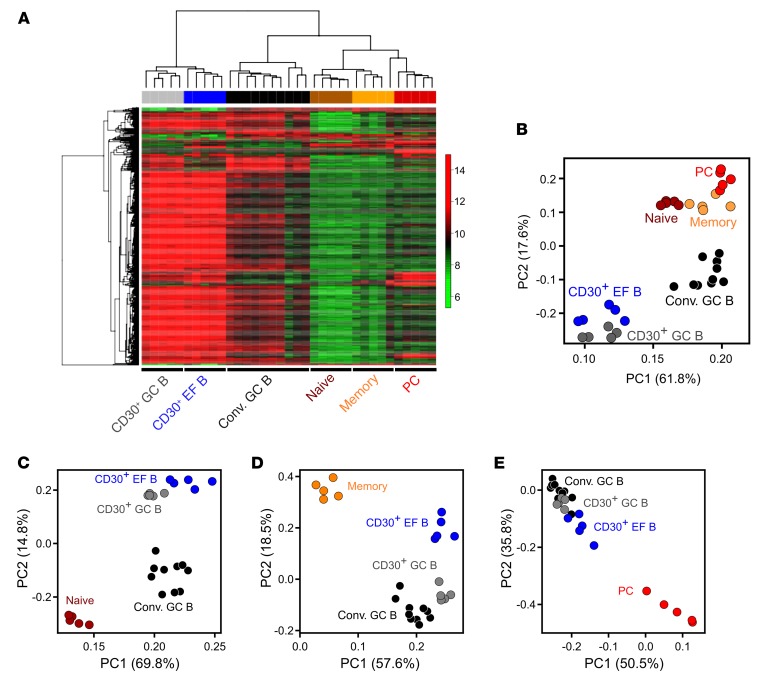
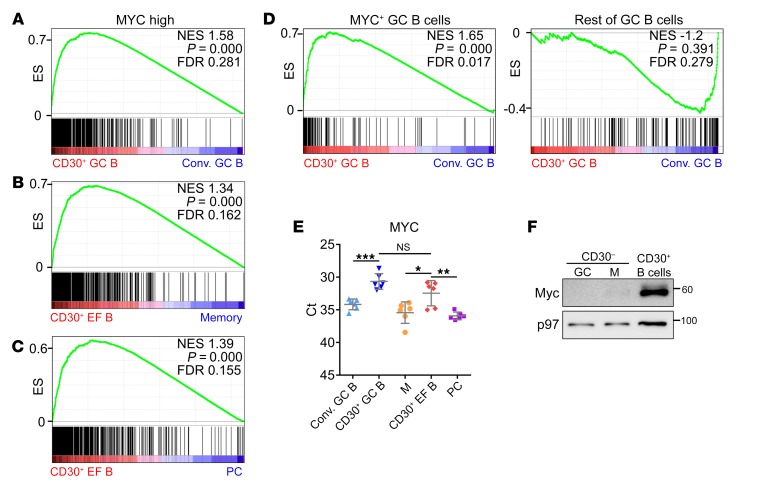
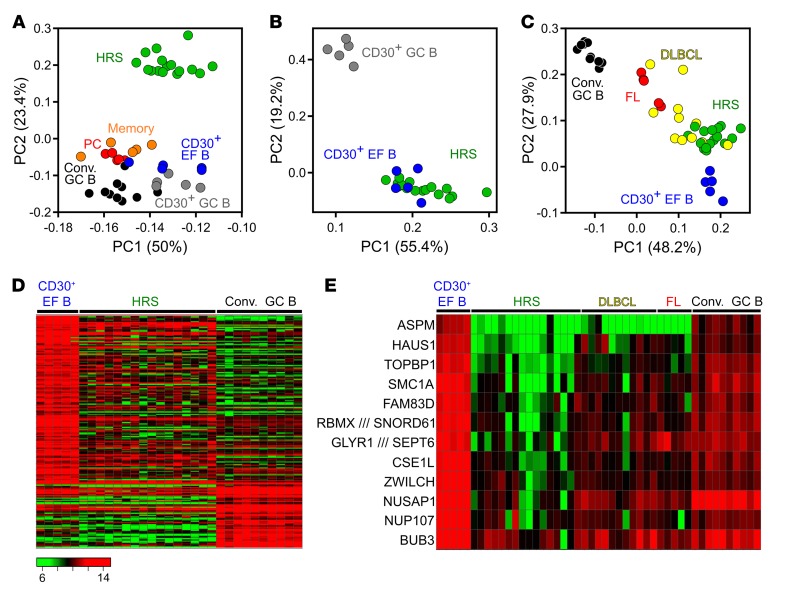
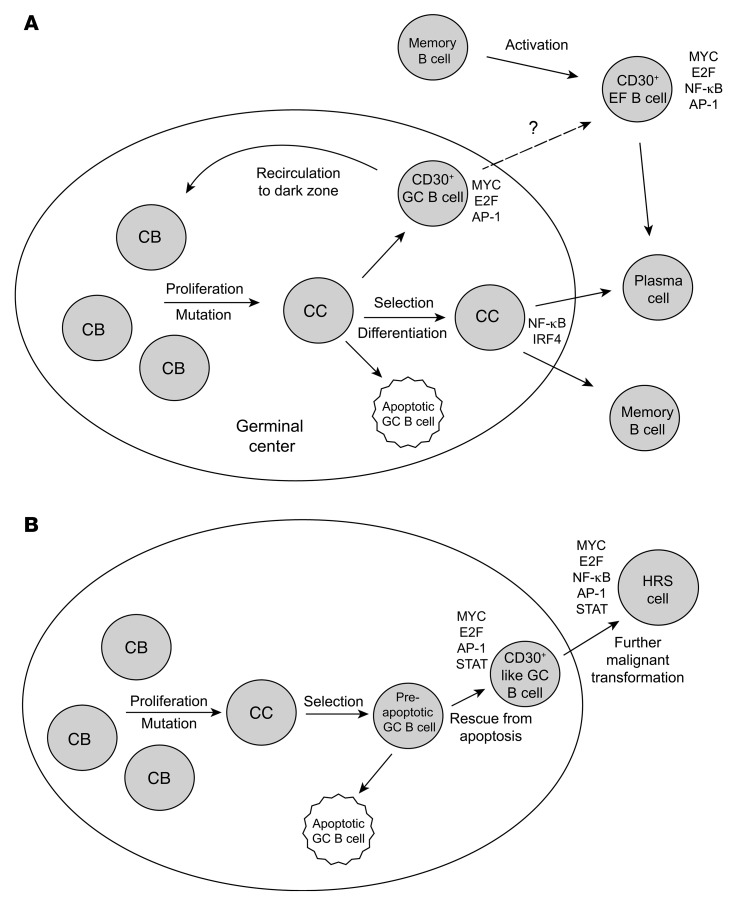
Very few B cells in germinal centers (GCs) and extrafollicular (EF) regions of lymph nodes express CD30. Their specific features and relationship to CD30-expressing Hodgkin and Reed/Sternberg (HRS) cells of Hodgkin lymphoma are unclear but highly relevant, because numerous patients with lymphoma are currently treated with an anti-CD30 immunotoxin. We performed a comprehensive analysis of human CD30+ B cells. Phenotypic and IgV gene analyses indicated that CD30+ GC B lymphocytes represent typical GC B cells, and that CD30+ EF B cells are mostly post-GC B cells. The transcriptomes of CD30+ GC and EF B cells largely overlapped, sharing a strong MYC signature, but were strikingly different from conventional GC B cells and memory B and plasma cells, respectively. CD30+ GC B cells represent MYC+ centrocytes redifferentiating into centroblasts; CD30+ EF B cells represent active, proliferating memory B cells. HRS cells shared typical transcriptome patterns with CD30+ B cells, suggesting that they originate from these lymphocytes or acquire their characteristic features during lymphomagenesis. By comparing HRS to normal CD30+ B cells we redefined aberrant and disease-specific features of HRS cells. A remarkable downregulation of genes regulating genomic stability and cytokinesis in HRS cells may explain their genomic instability and multinuclearity.
Keywords: B cells; Hematology; Hodgkins lymphoma; Immunology; Molecular pathology.
Conflict of interest statement
Figures
References
-
- Falini B, et al. CD30 (Ki-1) molecule: a new cytokine receptor of the tumor necrosis factor receptor superfamily as a tool for diagnosis and immunotherapy. Blood. 1995;85(1):1–14. - PubMed
-
- Stein H, et al. The expression of the Hodgkin’s disease associated antigen Ki-1 in reactive and neoplastic lymphoid tissue: evidence that Reed-Sternberg cells and histiocytic malignancies are derived from activated lymphoid cells. Blood. 1985;66(4):848–858. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
