

Aged murine hematopoietic stem cells drive aging-associated immune remodeling
- PMID: 29891535
- PMCID: PMC6137572
- DOI: 10.1182/blood-2018-02-831065
Aged murine hematopoietic stem cells drive aging-associated immune remodeling
Abstract
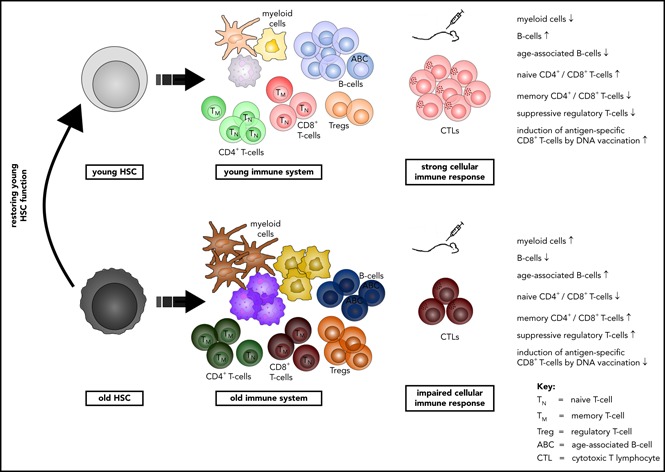
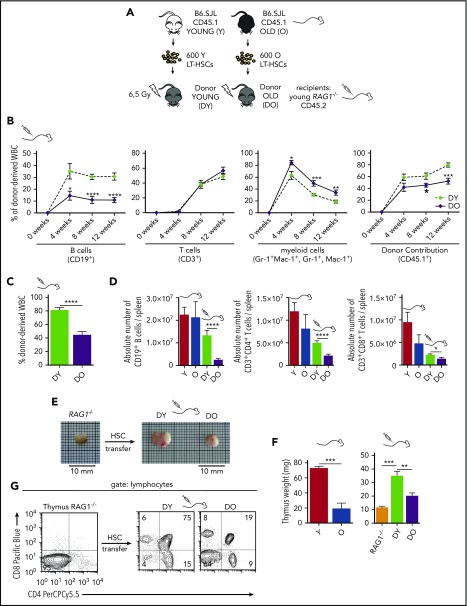
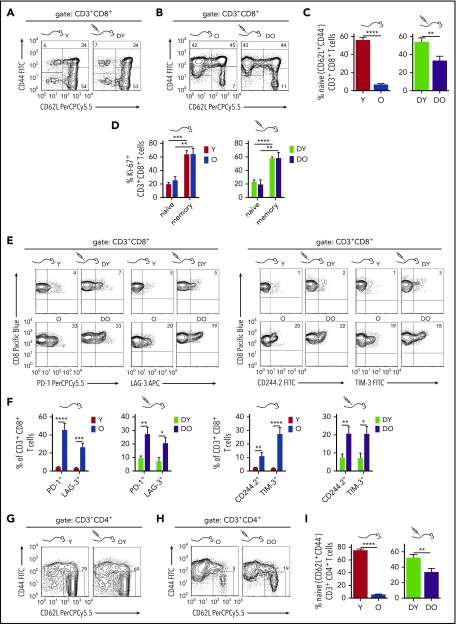
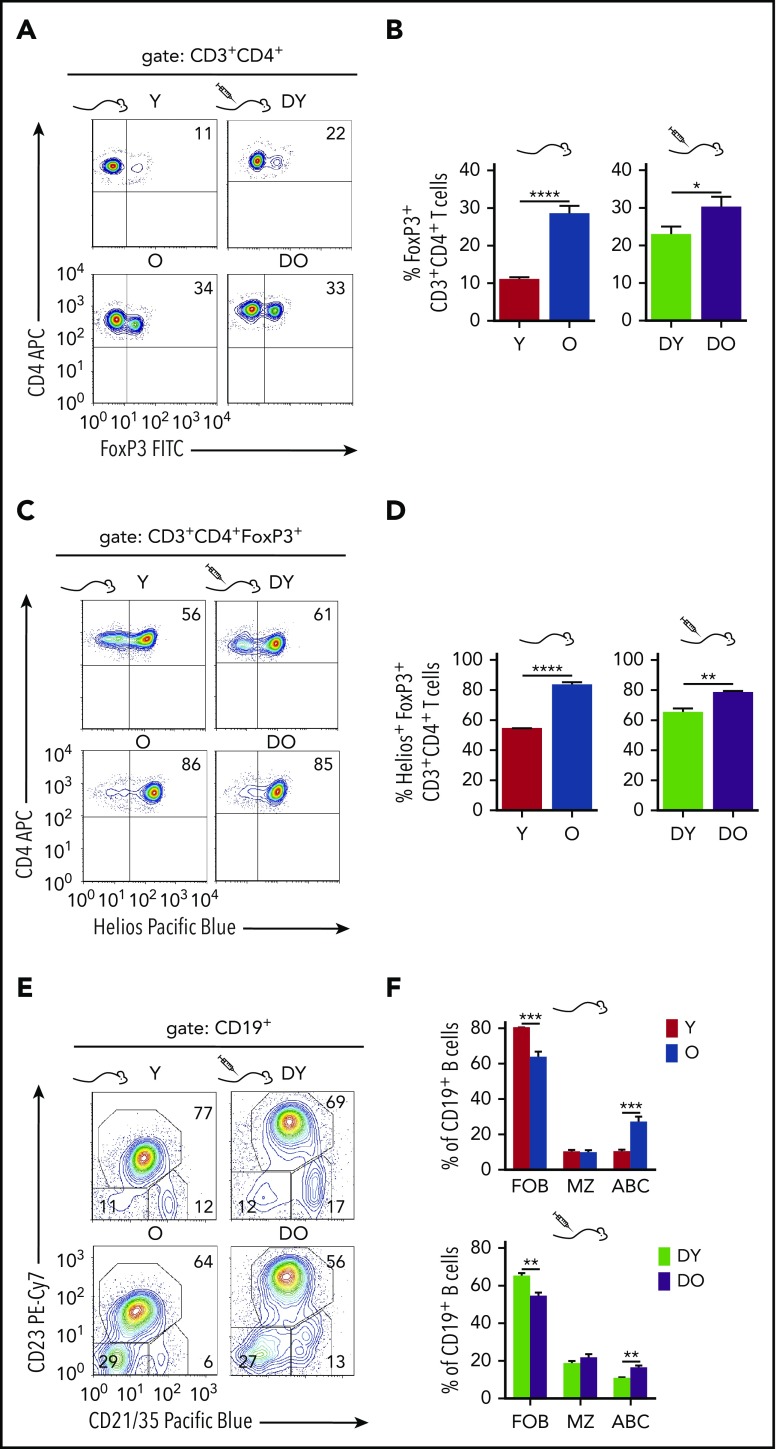
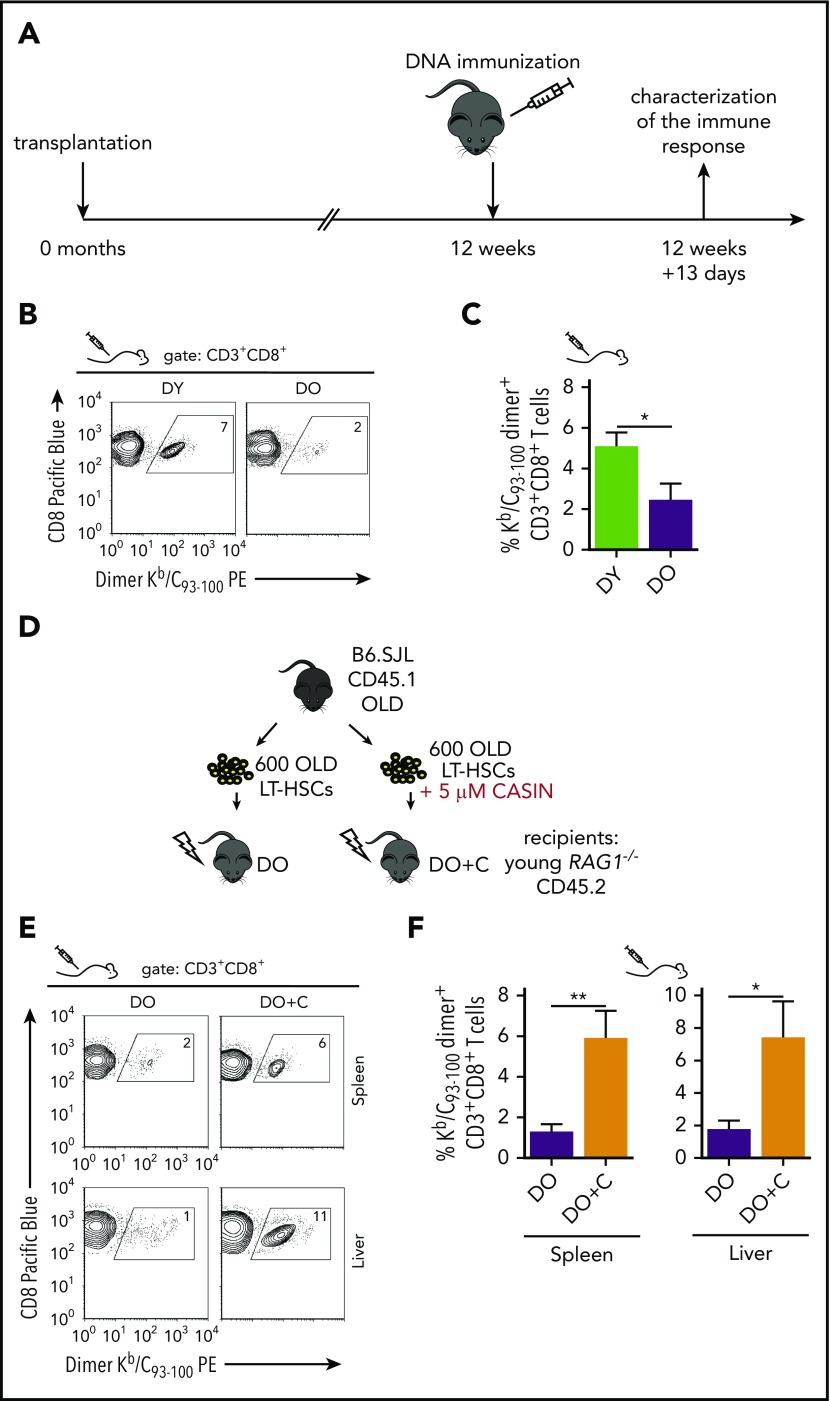
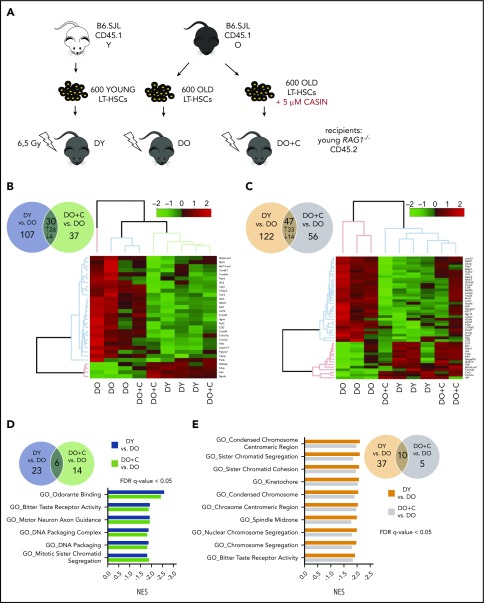
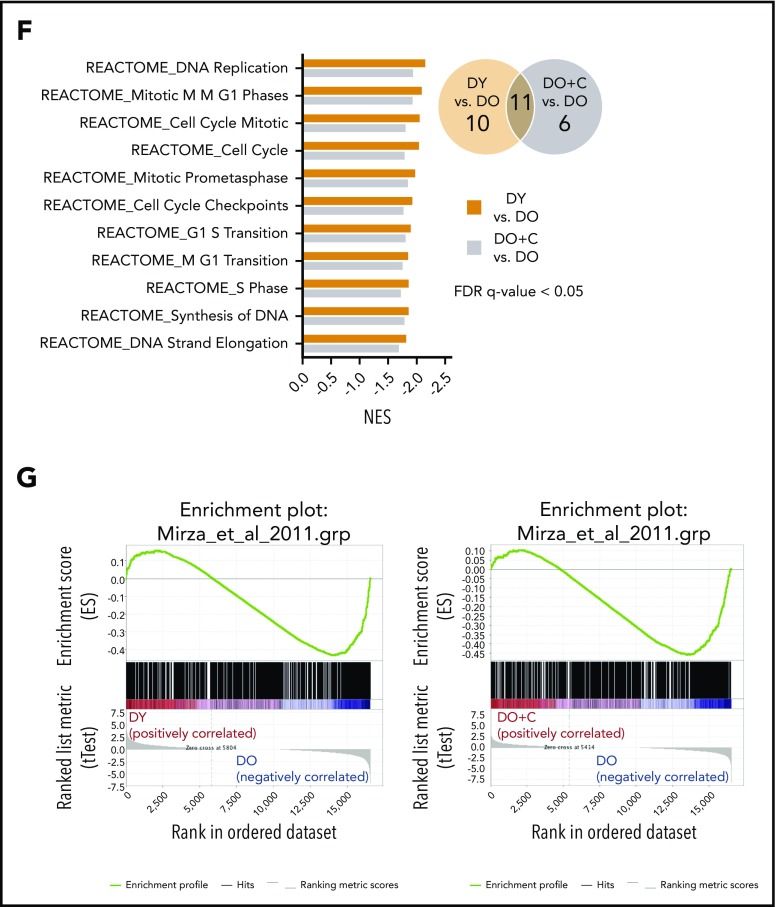
Aging-associated remodeling of the immune system impairs its functional integrity and contributes to increased morbidity and mortality in the elderly. Aging of hematopoietic stem cells (HSCs), from which all cells of the adaptive immune system ultimately originate, might play a crucial role in the remodeling of the aged immune system. We recently reported that aging of HSCs is, in part, driven by elevated activity of the small RhoGTPase Cdc42 and that aged HSCs can be rejuvenated in vitro by inhibition of the elevated Cdc42 activity in aged HSCs with the pharmacological compound CASIN. To study the quality of immune systems stemming selectively from young or aged HSCs, we established a HSC transplantation model in T- and B-cell-deficient young RAG1-/- hosts. We report that both phenotypic and functional changes in the immune system on aging are primarily a consequence of changes in the function of HSCs on aging and, to a large extent, independent of the thymus, as young and aged HSCs reconstituted distinct T- and B-cell subsets in RAG1-/- hosts that mirrored young and aged immune systems. Importantly, aged HSCs treated with CASIN reestablished an immune system similar to that of young animals, and thus capable of mounting a strong immune response to vaccination. Our studies further imply that epigenetic signatures already imprinted in aged HSCs determine the transcriptional profile and function of HSC-derived T and B cells.
© 2018 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures
Comment in
-
CASIN the joint: immune aging at the stem cell level.Blood. 2018 Aug 9;132(6):553-554. doi: 10.1182/blood-2018-06-858696. Blood. 2018. PMID: 30093381 No abstract available.
References
-
- Black S, De Gregorio E, Rappuoli R. Developing vaccines for an aging population. Sci Transl Med. 2015;7(281):281ps8. - PubMed
-
- Boraschi D, Aguado MT, Dutel C, et al. The gracefully aging immune system. Sci Transl Med. 2013;5(185):185ps8. - PubMed
-
- Nikolich-Žugich J. The twilight of immunity: emerging concepts in aging of the immune system. Nat Immunol. 2018;19(1):10-19. - PubMed
-
- Aw D, Silva AB, Maddick M, von Zglinicki T, Palmer DB. Architectural changes in the thymus of aging mice. Aging Cell. 2008;7(2):158-167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
