

Single-Molecule Fluorescence Applied to Translation
- PMID: 29891562
- PMCID: PMC6314065
- DOI: 10.1101/cshperspect.a032714
Single-Molecule Fluorescence Applied to Translation
Abstract
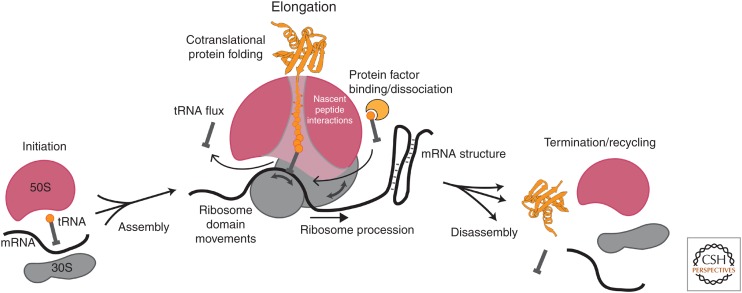
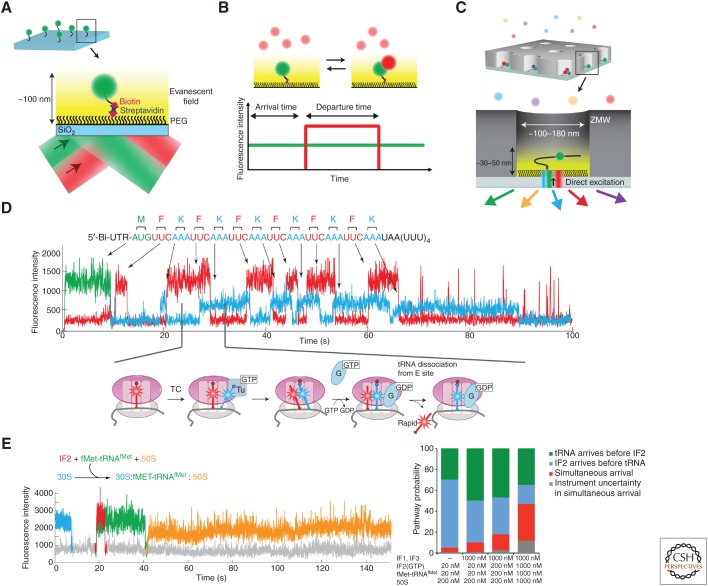
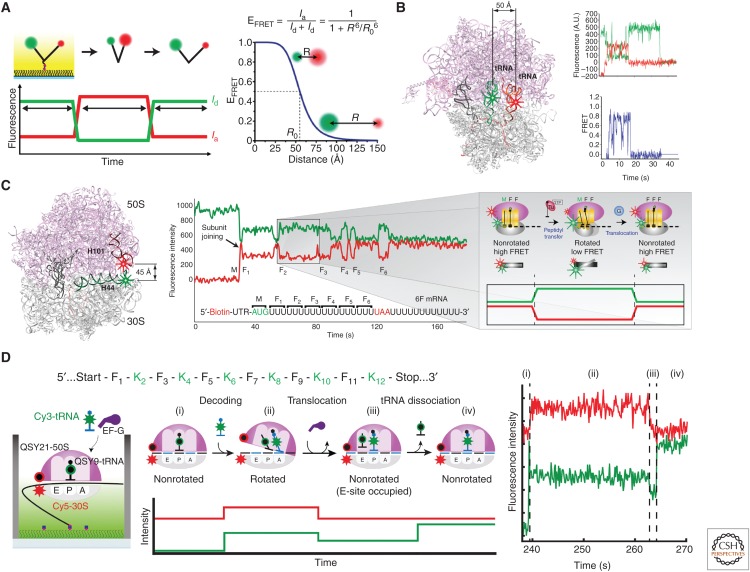
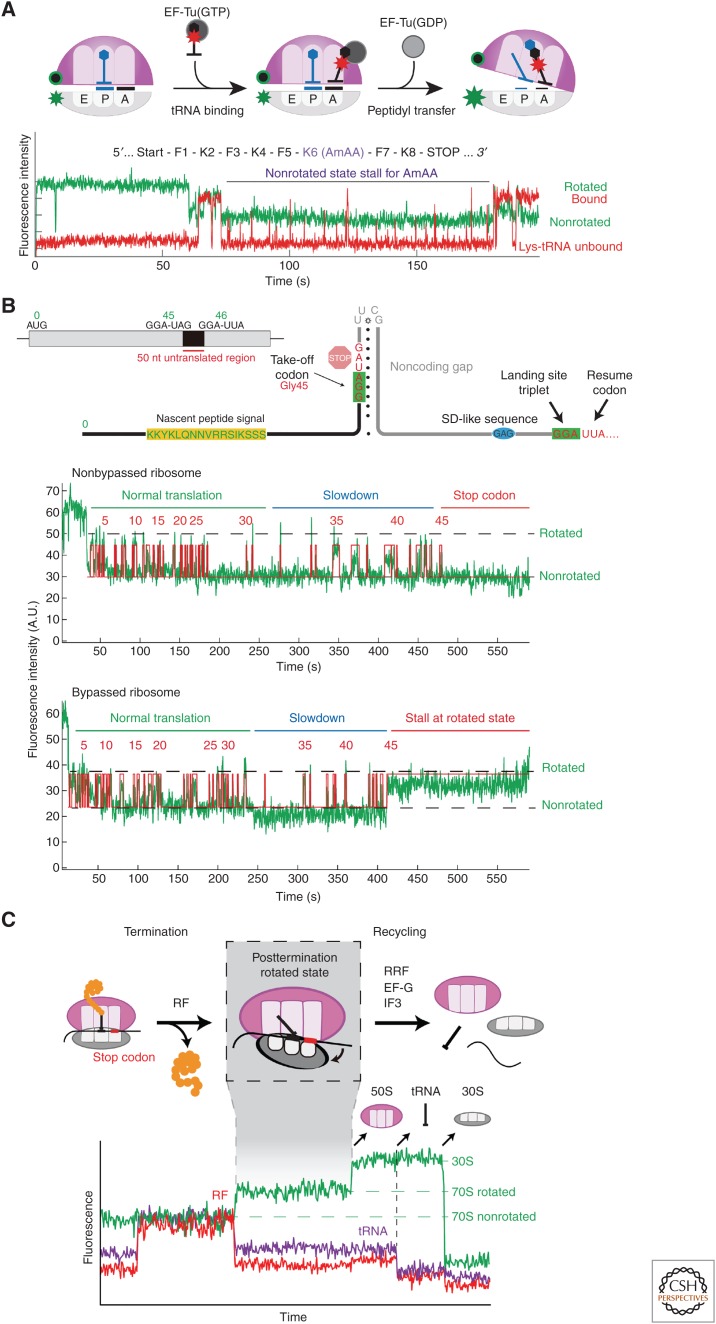
Single-molecule fluorescence methods have illuminated the dynamics of the translational machinery. Structural and bulk biochemical experiments have provided detailed atomic and global mechanistic views of translation, respectively. Single-molecule studies of translation have bridged these views by temporally connecting the conformational and compositional states defined from structural data within the mechanistic framework of translation produced from biochemical studies. Here, we discuss the context for applying different single-molecule fluorescence experiments, and present recent applications to studying prokaryotic and eukaryotic translation. We underscore the power of observing single translating ribosomes to delineate and sort complex mechanistic pathways during initiation and elongation, and discuss future applications of current and improved technologies.
Copyright © 2019 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA. 2000. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 289: 905–920. - PubMed
-
- Barbet-Massin E, Huang CT, Daebel V, Hsu ST, Reif B. 2015. Site-specific solid-state NMR studies of “trigger factor” in complex with the large ribosomal subunit 50S. Angew Chem Int Ed Engl 54: 4367–4369. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources