

NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum
- PMID: 29891874
- PMCID: PMC5995848
- DOI: 10.1038/s41598-018-27274-8
NtLTP4, a lipid transfer protein that enhances salt and drought stresses tolerance in Nicotiana tabacum
Abstract
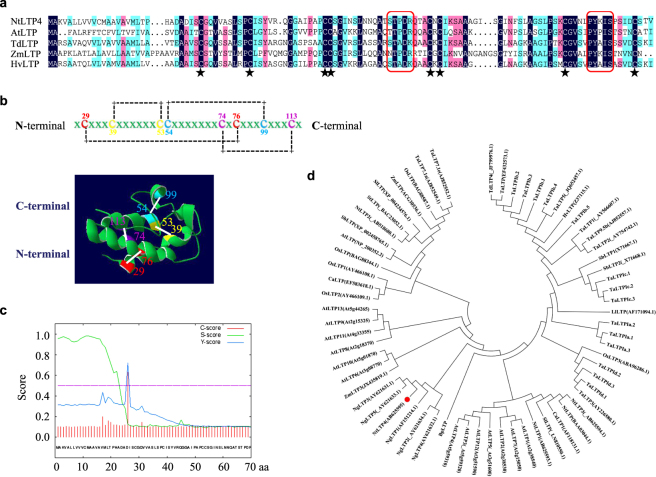
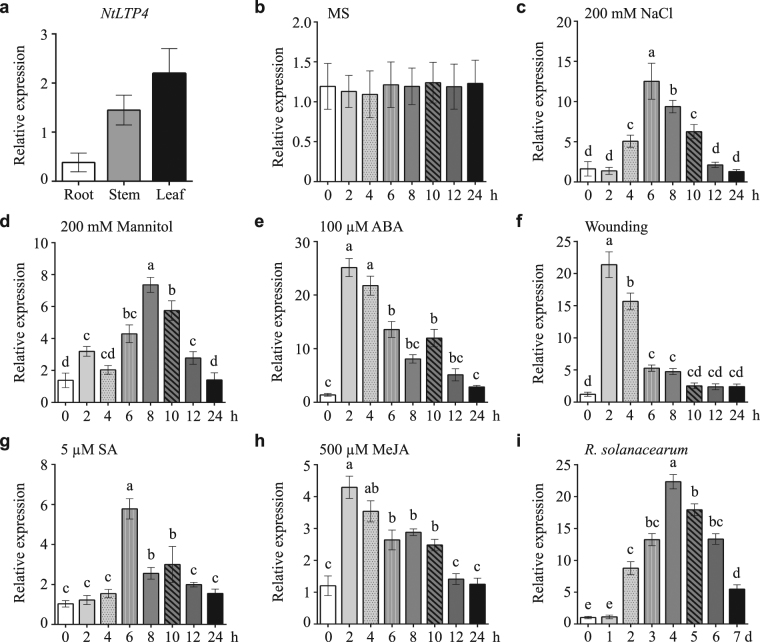
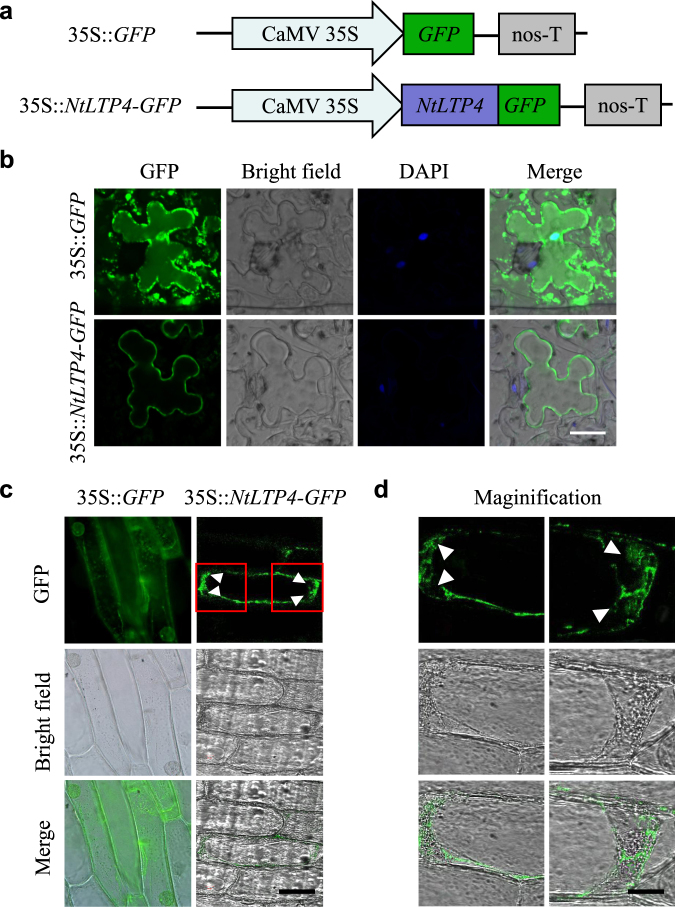
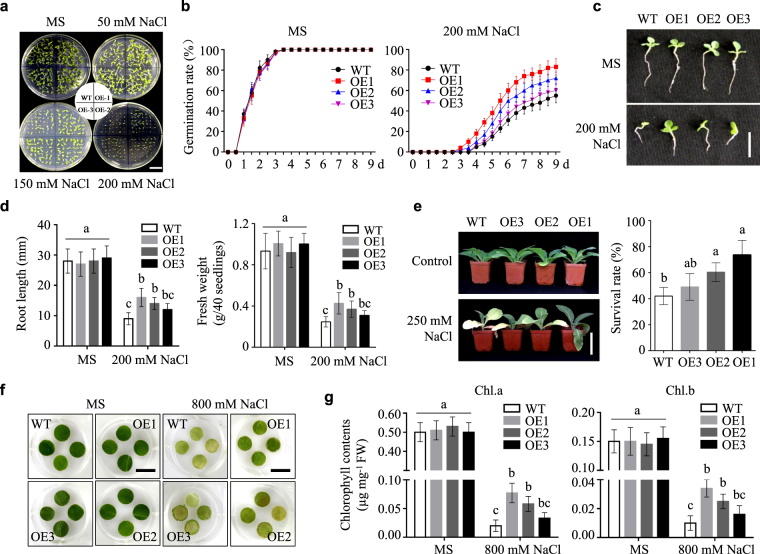
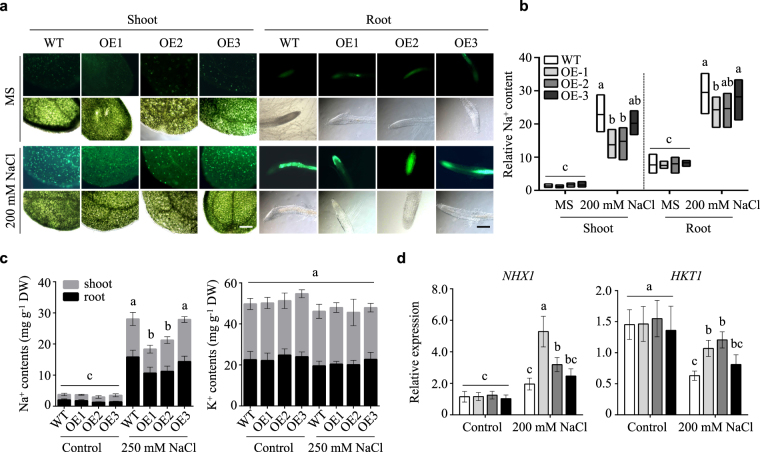
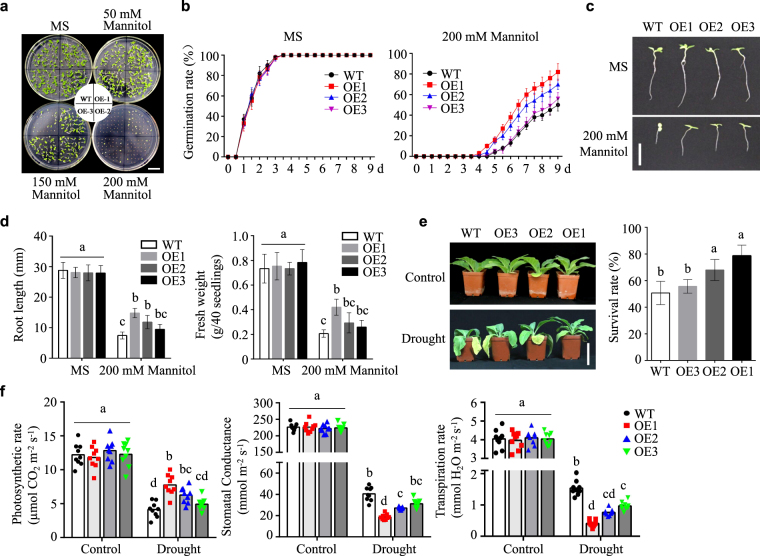
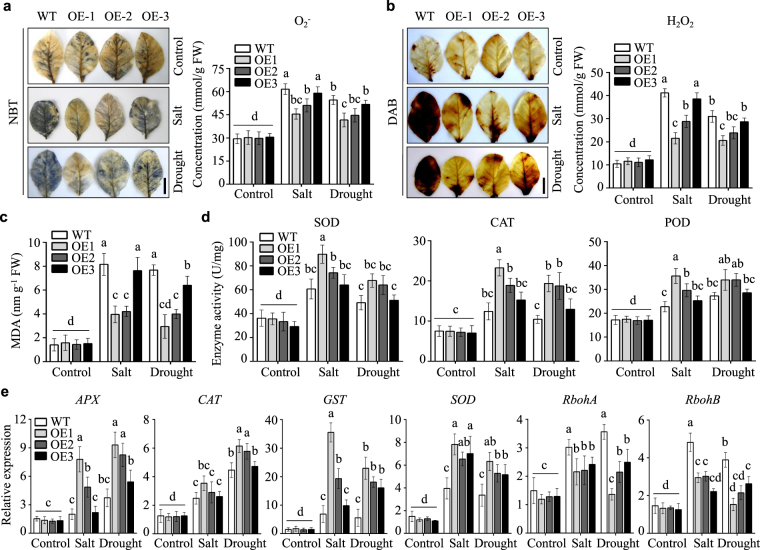
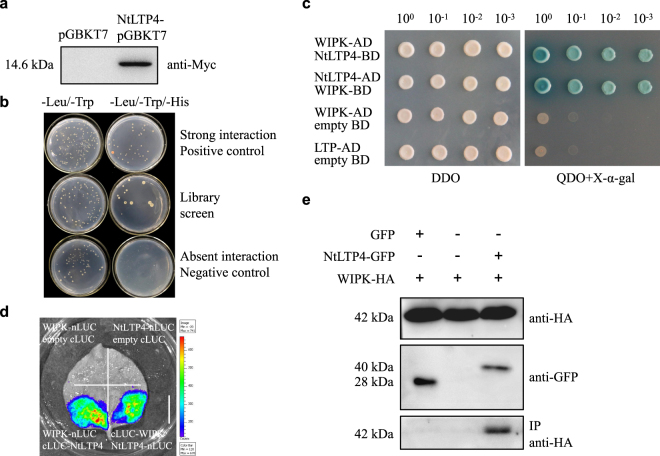
Lipid transfer proteins (LTPs), a class of small, ubiquitous proteins, play critical roles in various environmental stresses. However, their precise biological functions remain unknown. Here we isolated an extracellular matrix-localised LTP, NtLTP4, from Nicotiana tabacum. The overexpression of NtLTP4 in N. tabacum enhanced resistance to salt and drought stresses. Upon exposure to high salinity, NtLTP4-overexpressing lines (OE lines) accumulated low Na+ levels. Salt-responsive genes, including Na+/H+ exchangers (NHX1) and high-affinity K+ transporter1 (HKT1), were dramatically higher in OE lines than in wild-type lines. NtLTP4 might regulate transcription levels of NHX1 and HKT1 to alleviate the toxicity of Na+. Interestingly, OE lines enhanced the tolerance of N. tabacum to drought stress by reducing the transpiration rate. Moreover, NtLTP4 could increase reactive oxygen species (ROS)-scavenging enzyme activity and expression levels to scavenge excess ROS under drought and high salinity conditions. We used a two-hybrid yeast system and screened seven putative proteins that interact with NtLTP4 in tobacco. An MAPK member, wound-induced protein kinase, was confirmed to interact with NtLTP4 via co-immunoprecipitation and a firefly luciferase complementation imaging assay. Taken together, this is the first functional analysis of NtLTP4, and proves that NtLTP4 positively regulates salt and drought stresses in N. tabacum.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Li, P. et al. TheA UDP-glycosyltransferases UGT79B2 and 79B3, contribute to cold, salt and drought stress tolerance via modulating anthocyanin accumulation. The Plant journal: for cell and molecular biology, 10.1111/tpj.13324 (2016). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
