

An interactome perturbation framework prioritizes damaging missense mutations for developmental disorders
- PMID: 29892012
- PMCID: PMC6314957
- DOI: 10.1038/s41588-018-0130-z
An interactome perturbation framework prioritizes damaging missense mutations for developmental disorders
Abstract
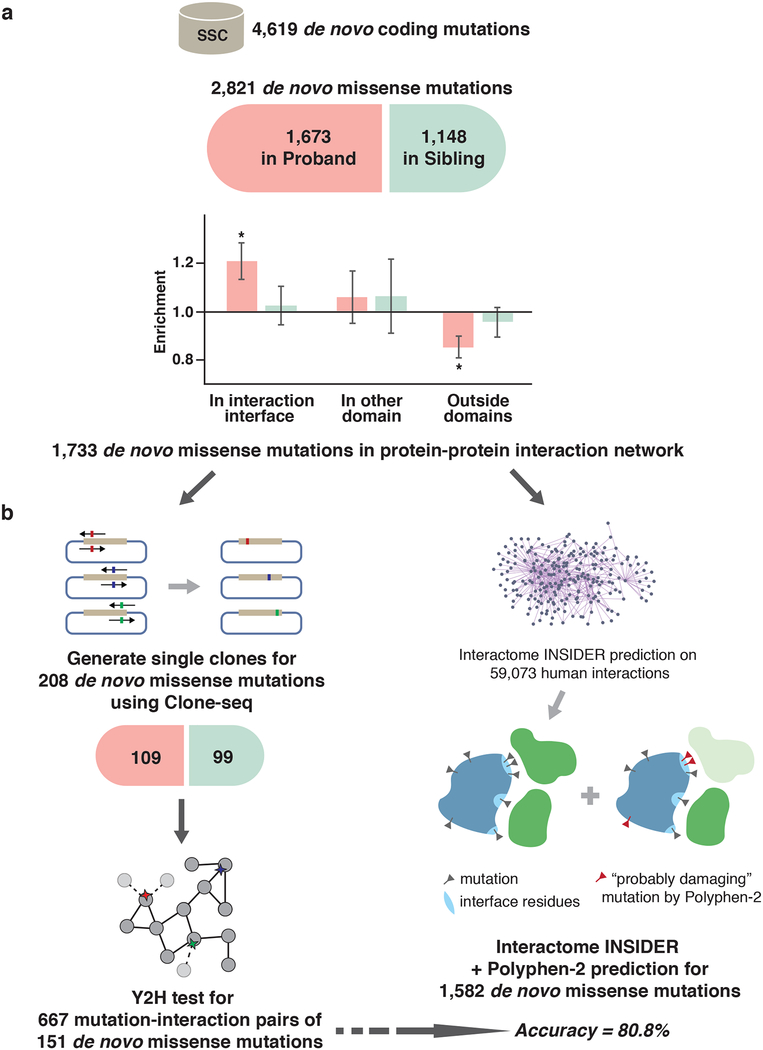
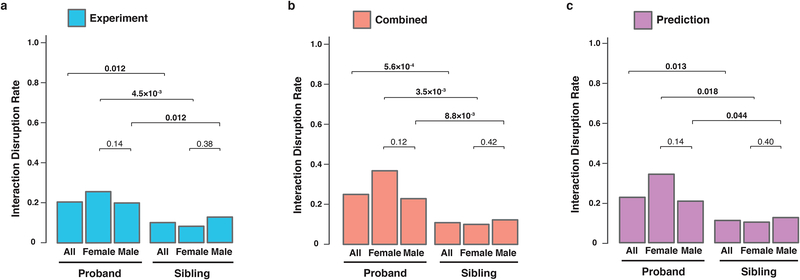
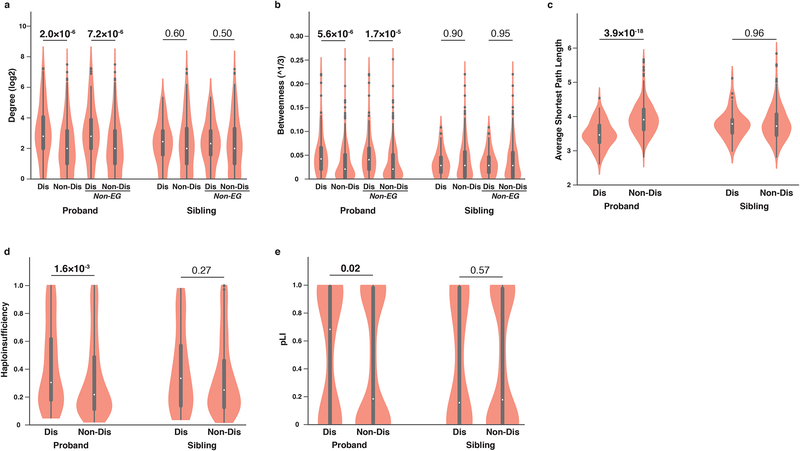
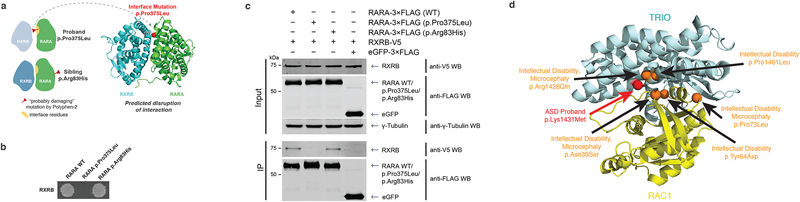
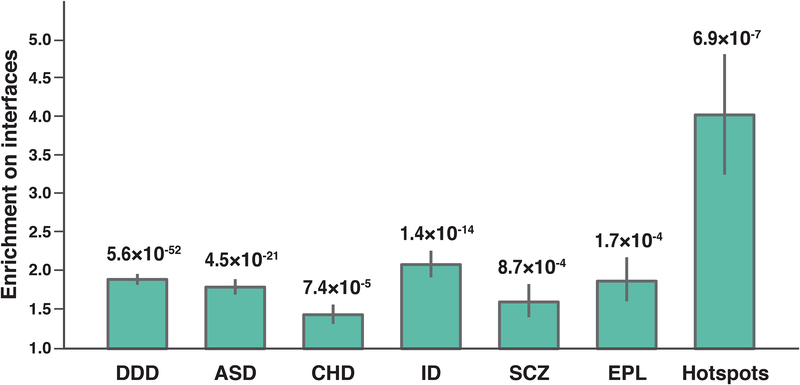
Identifying disease-associated missense mutations remains a challenge, especially in large-scale sequencing studies. Here we establish an experimentally and computationally integrated approach to investigate the functional impact of missense mutations in the context of the human interactome network and test our approach by analyzing ~2,000 de novo missense mutations found in autism subjects and their unaffected siblings. Interaction-disrupting de novo missense mutations are more common in autism probands, principally affect hub proteins, and disrupt a significantly higher fraction of hub interactions than in unaffected siblings. Moreover, they tend to disrupt interactions involving genes previously implicated in autism, providing complementary evidence that strengthens previously identified associations and enhances the discovery of new ones. Importantly, by analyzing de novo missense mutation data from six disorders, we demonstrate that our interactome perturbation approach offers a generalizable framework for identifying and prioritizing missense mutations that contribute to the risk of human disease.
Conflict of interest statement
Competing Interests
The authors declare no competing financial interests.
Figures
References
-
- Ropers HH Genetics of early onset cognitive impairment. Annu Rev Genomics Hum Genet 11, 161–87 (2010). - PubMed
-
- Devlin B & Scherer SW Genetic architecture in autism spectrum disorder. Curr Opin Genet Dev 22, 229–37 (2012). - PubMed
-
- Bruneau BG The developmental genetics of congenital heart disease. Nature 451, 943–8 (2008). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
