

Impaired hippocampal representation of place in the Fmr1-knockout mouse model of fragile X syndrome
- PMID: 29892074
- PMCID: PMC5995880
- DOI: 10.1038/s41598-018-26853-z
Impaired hippocampal representation of place in the Fmr1-knockout mouse model of fragile X syndrome
Abstract
Fragile X syndrome (FXS) is an X-chromosome linked intellectual disability and the most common known inherited single gene cause of autism spectrum disorder (ASD). Building upon demonstrated deficits in neuronal plasticity and spatial memory in FXS, we investigated how spatial information processing is affected in vivo in an FXS mouse model (Fmr1-KO). Healthy hippocampal neurons (so-called place cells) exhibit place-related activity during spatial exploration, and their firing fields tend to remain stable over time. In contrast, we find impaired stability and reduced specificity of Fmr1-KO spatial representations. This is a potential biomarker for the cognitive dysfunction observed in FXS, informative on the ability to integrate sensory information into an abstract representation and successfully retain this conceptual memory. Our results provide key insight into the biological mechanisms underlying cognitive disabilities in FXS and ASD, paving the way for a targeted approach to remedy these.
Conflict of interest statement
The authors declare no competing interests.
Figures
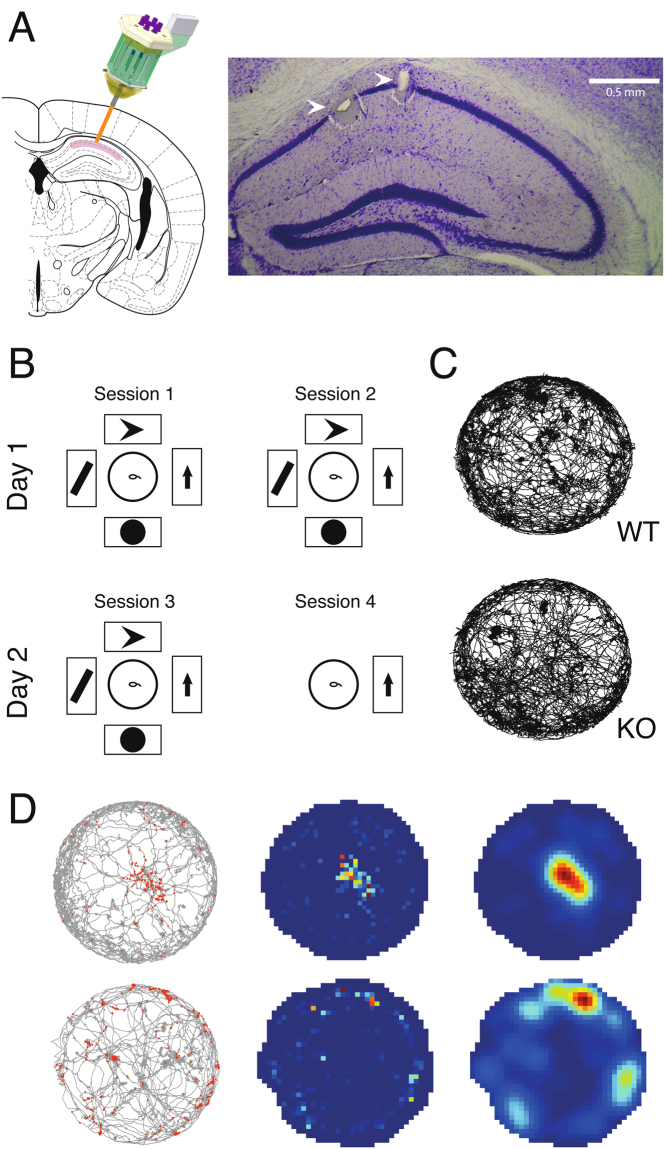
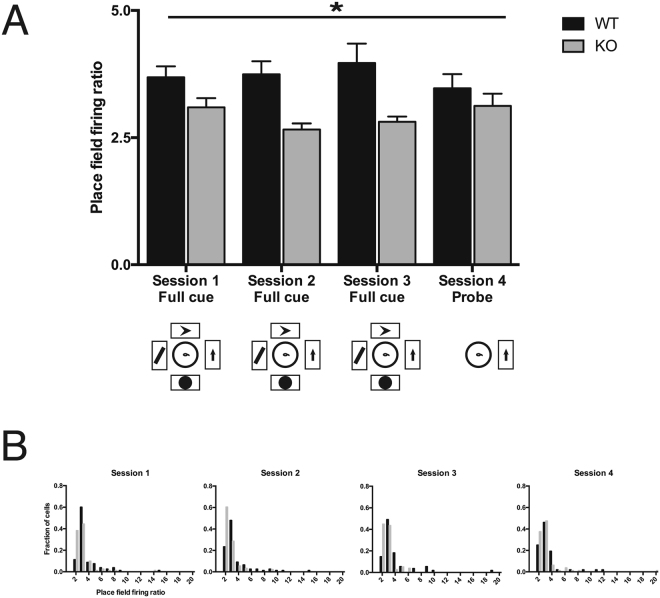
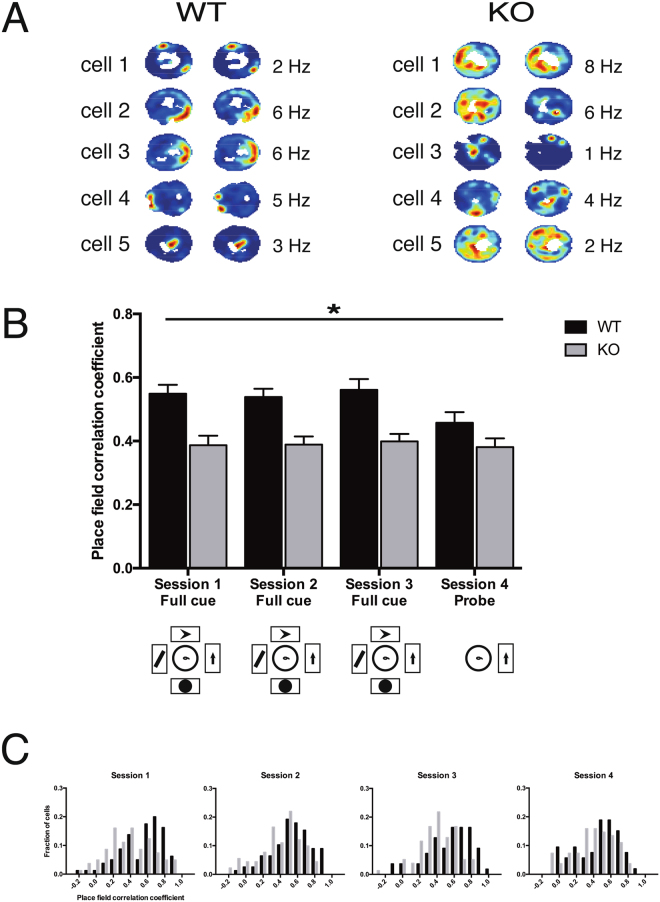
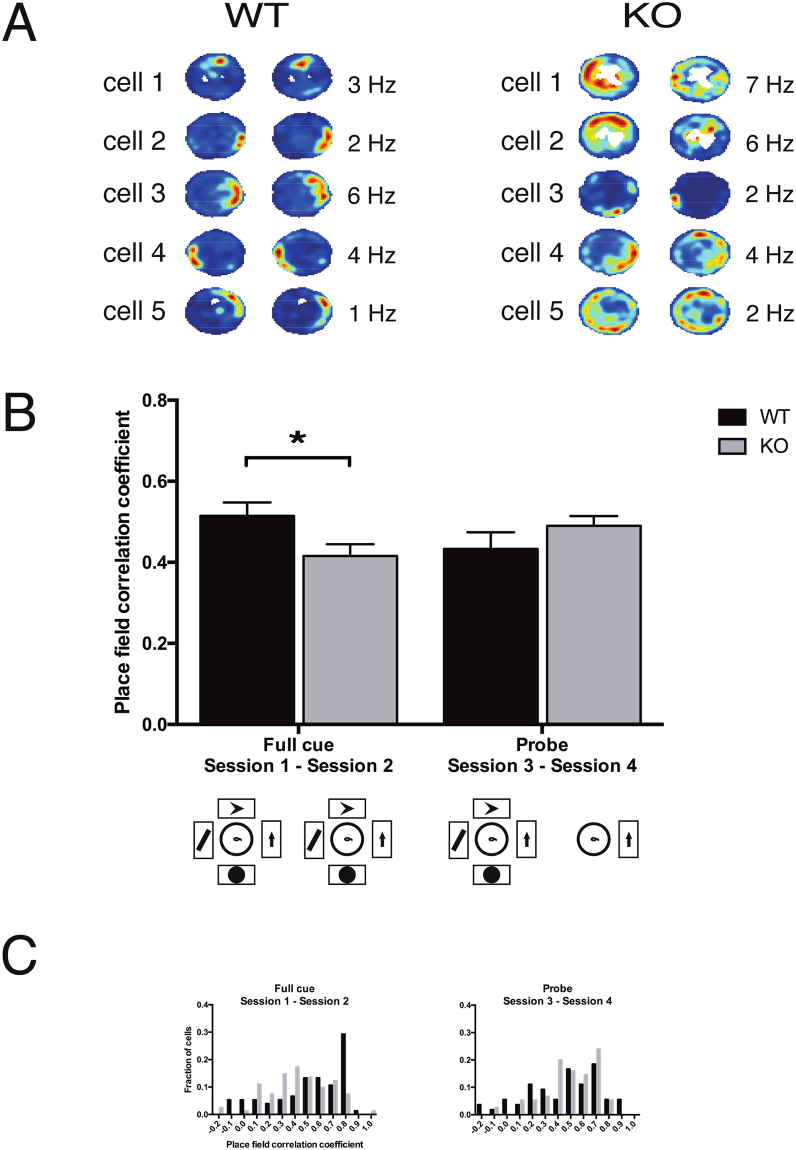
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
