

Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration
- PMID: 29894310
- PMCID: PMC6063471
- DOI: 10.1172/JCI120972
Purinergic P2X4 receptors and mitochondrial ATP production regulate T cell migration
Abstract
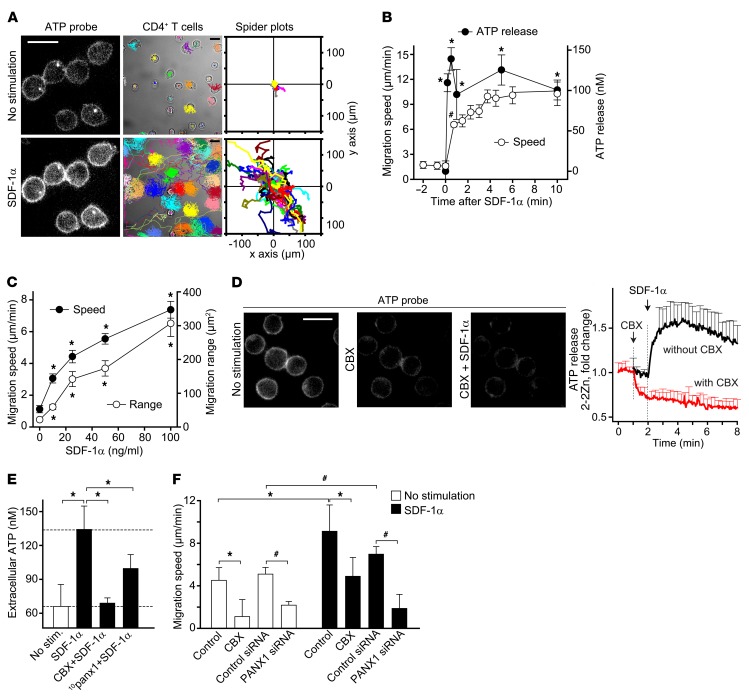
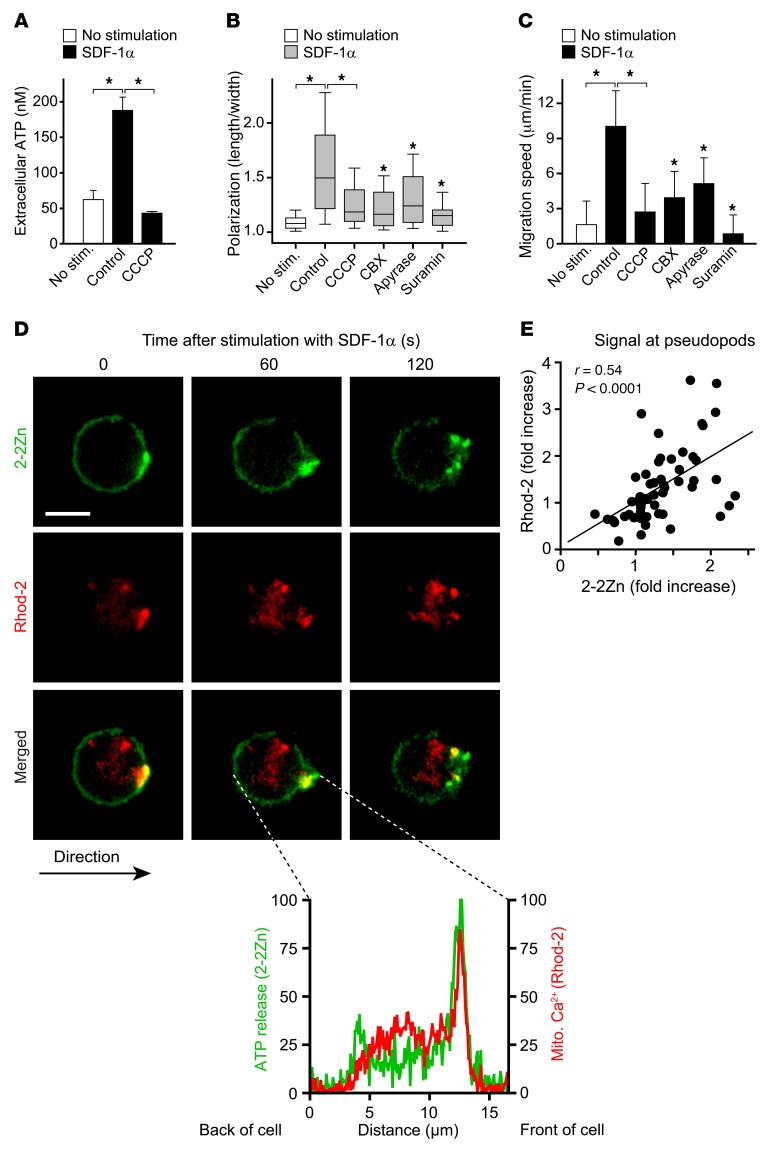
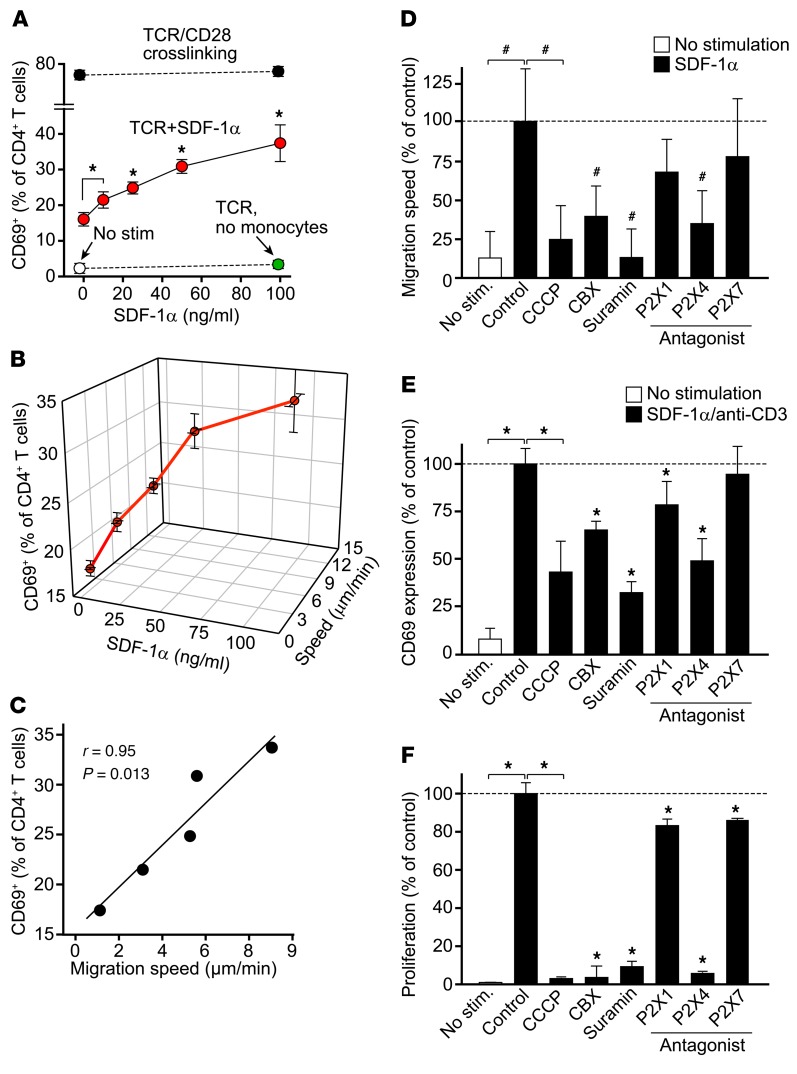
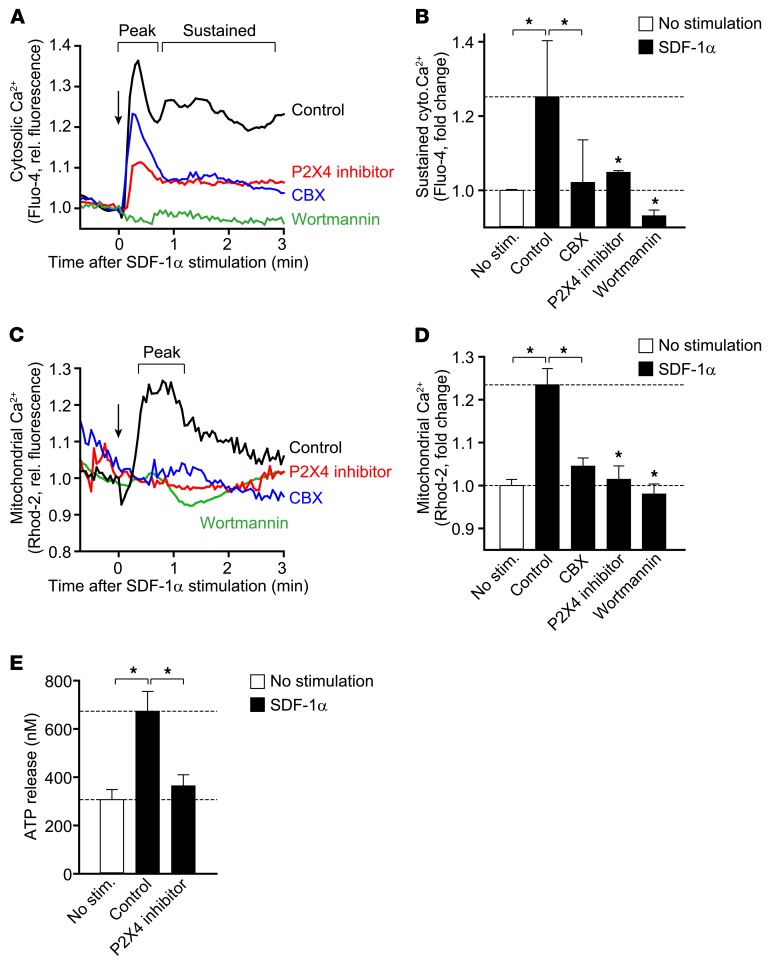
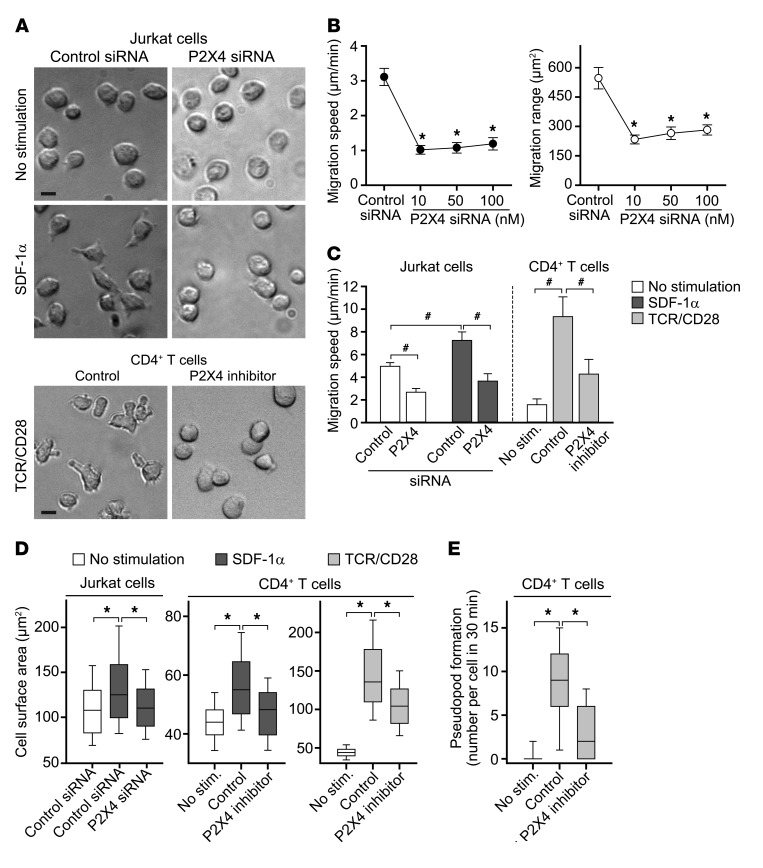
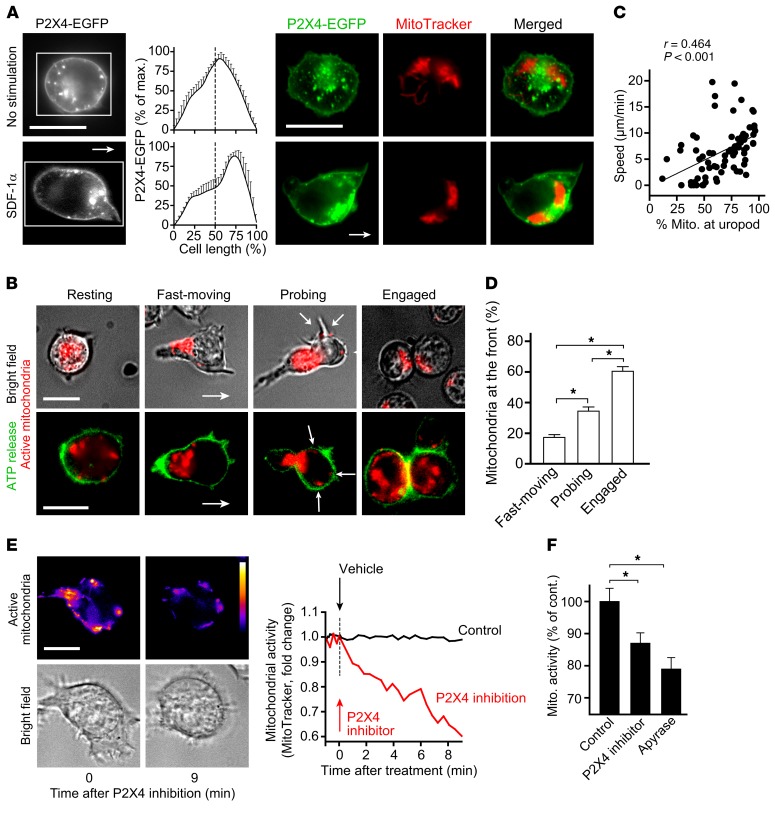
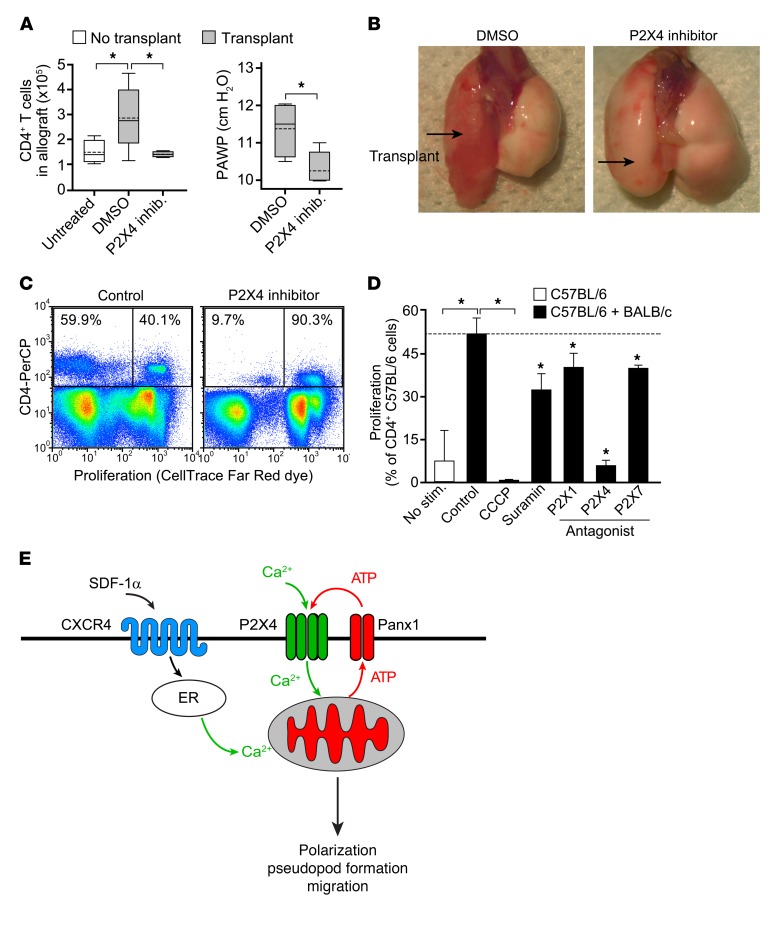
T cells must migrate in order to encounter antigen-presenting cells (APCs) and to execute their varied functions in immune defense and inflammation. ATP release and autocrine signaling through purinergic receptors contribute to T cell activation at the immune synapse that T cells form with APCs. Here, we show that T cells also require ATP release and purinergic signaling for their migration to APCs. We found that the chemokine stromal-derived factor-1α (SDF-1α) triggered mitochondrial ATP production, rapid bursts of ATP release, and increased migration of primary human CD4+ T cells. This process depended on pannexin-1 ATP release channels and autocrine stimulation of P2X4 receptors. SDF-1α stimulation caused localized accumulation of mitochondria with P2X4 receptors near the front of cells, resulting in a feed-forward signaling mechanism that promotes cellular Ca2+ influx and sustains mitochondrial ATP synthesis at levels needed for pseudopod protrusion, T cell polarization, and cell migration. Inhibition of P2X4 receptors blocked the activation and migration of T cells in vitro. In a mouse lung transplant model, P2X4 receptor antagonist treatment prevented the recruitment of T cells into allograft tissue and the rejection of lung transplants. Our findings suggest that P2X4 receptors are therapeutic targets for immunomodulation in transplantation and inflammatory diseases.
Keywords: Cell Biology; Mitochondria; T cells; Transplantation.
Conflict of interest statement
Figures
Similar articles
-
Pannexin-1 hemichannel-mediated ATP release together with P2X1 and P2X4 receptors regulate T-cell activation at the immune synapse.Blood. 2010 Nov 4;116(18):3475-84. doi: 10.1182/blood-2010-04-277707. Epub 2010 Jul 21. Blood. 2010. PMID: 20660288 Free PMC article.
-
Frontline Science: P2Y11 receptors support T cell activation by directing mitochondrial trafficking to the immune synapse.J Leukoc Biol. 2021 Mar;109(3):497-508. doi: 10.1002/JLB.2HI0520-191R. Epub 2020 Jun 12. J Leukoc Biol. 2021. PMID: 32531829 Free PMC article.
-
The purinergic receptor P2Y11 choreographs the polarization, mitochondrial metabolism, and migration of T lymphocytes.Sci Signal. 2020 Sep 29;13(651):eaba3300. doi: 10.1126/scisignal.aba3300. Sci Signal. 2020. PMID: 32994212 Free PMC article.
-
Mitochondria Synergize With P2 Receptors to Regulate Human T Cell Function.Front Immunol. 2020 Sep 29;11:549889. doi: 10.3389/fimmu.2020.549889. eCollection 2020. Front Immunol. 2020. PMID: 33133068 Free PMC article. Review.
-
Contribution of P2X4 Receptors to CNS Function and Pathophysiology.Int J Mol Sci. 2020 Aug 3;21(15):5562. doi: 10.3390/ijms21155562. Int J Mol Sci. 2020. PMID: 32756482 Free PMC article. Review.
Cited by
-
Ivermectin converts cold tumors hot and synergizes with immune checkpoint blockade for treatment of breast cancer.NPJ Breast Cancer. 2021 Mar 2;7(1):22. doi: 10.1038/s41523-021-00229-5. NPJ Breast Cancer. 2021. PMID: 33654071 Free PMC article.
-
Blocking pannexin1 reduces airway inflammation in a murine model of asthma.Am J Transl Res. 2020 Jul 15;12(7):4074-4083. eCollection 2020. Am J Transl Res. 2020. PMID: 32774761 Free PMC article.
-
Metabolic adaptation of lymphocytes in immunity and disease.Immunity. 2022 Jan 11;55(1):14-30. doi: 10.1016/j.immuni.2021.12.012. Immunity. 2022. PMID: 35021054 Free PMC article. Review.
-
Airway brush cells generate cysteinyl leukotrienes through the ATP sensor P2Y2.Sci Immunol. 2020 Jan 17;5(43):eaax7224. doi: 10.1126/sciimmunol.aax7224. Epub 2020 Jan 17. Sci Immunol. 2020. PMID: 31953256 Free PMC article.
-
Macrophage-P2X4 receptors pathway is essential to persistent inflammatory muscle hyperalgesia onset, and is prevented by physical exercise.PLoS One. 2025 Feb 11;20(2):e0318107. doi: 10.1371/journal.pone.0318107. eCollection 2025. PLoS One. 2025. PMID: 39932994 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
