

Structure and Composition Define Immunorecognition of Nucleic Acid Nanoparticles
- PMID: 29894623
- PMCID: PMC6540121
- DOI: 10.1021/acs.nanolett.8b01283
Structure and Composition Define Immunorecognition of Nucleic Acid Nanoparticles
Abstract
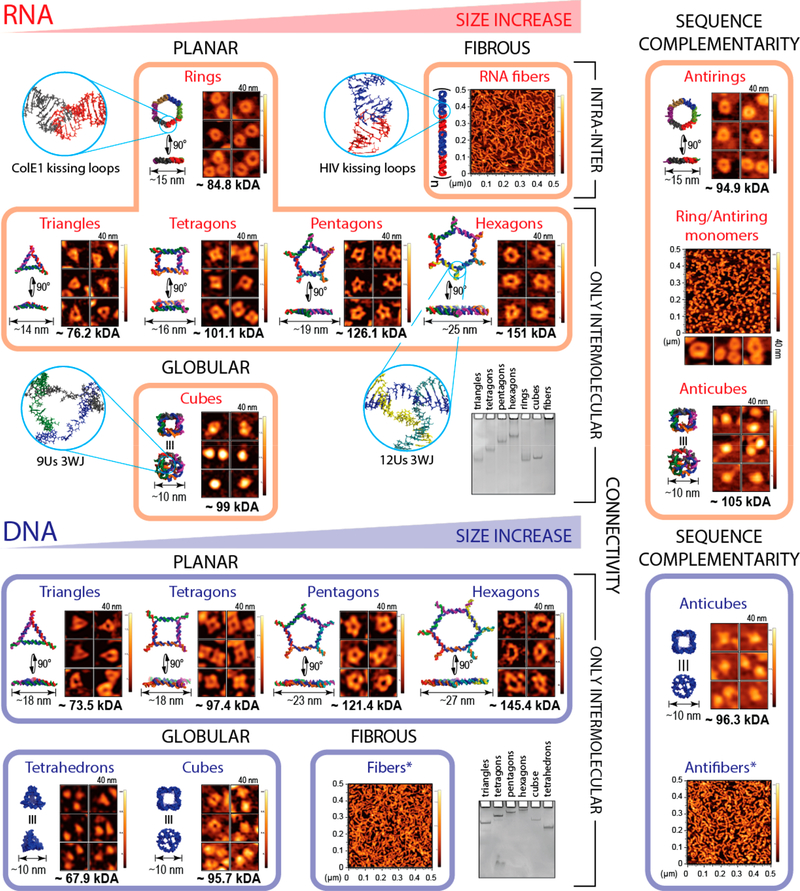
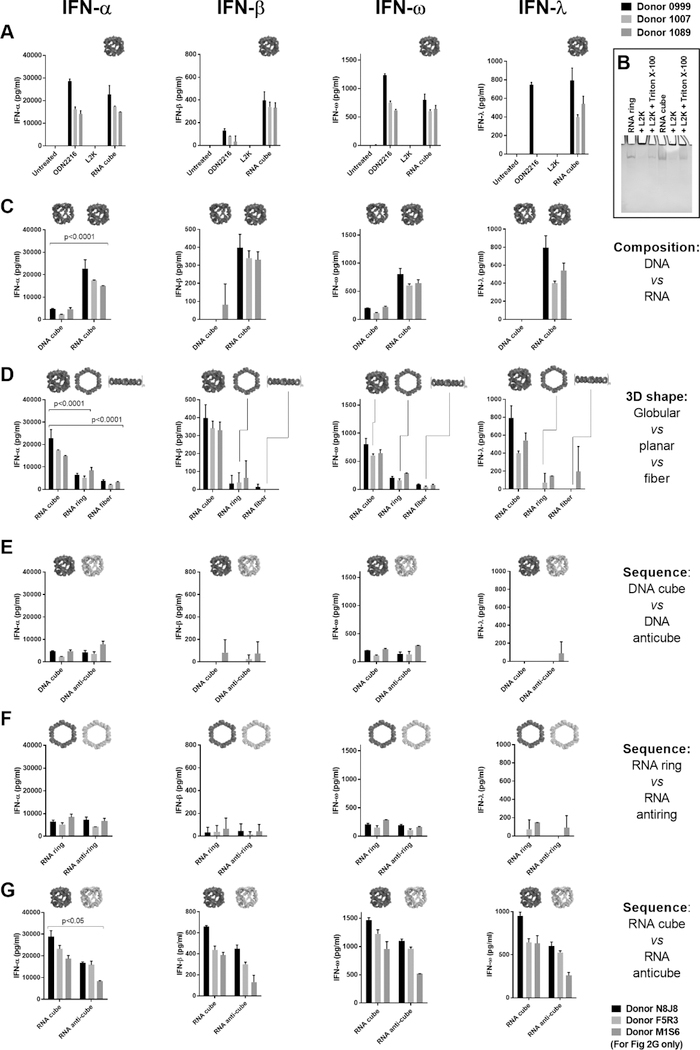
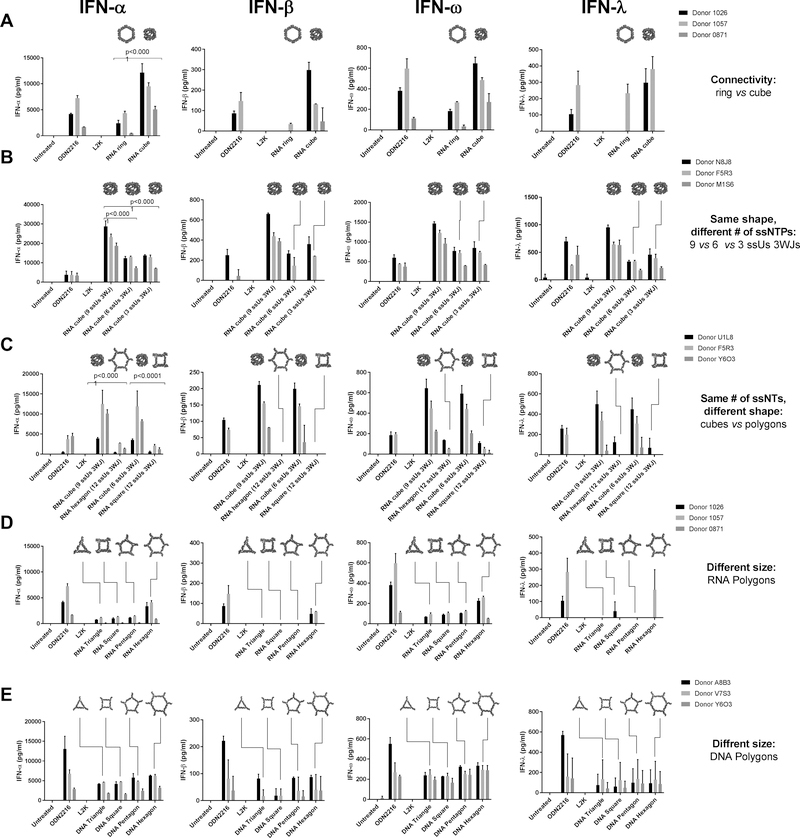
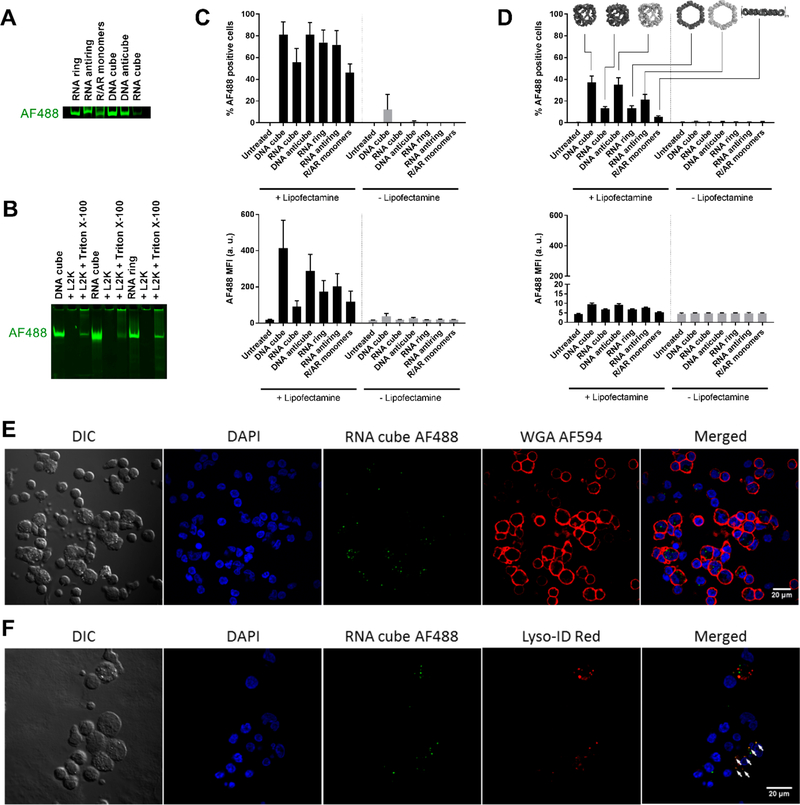
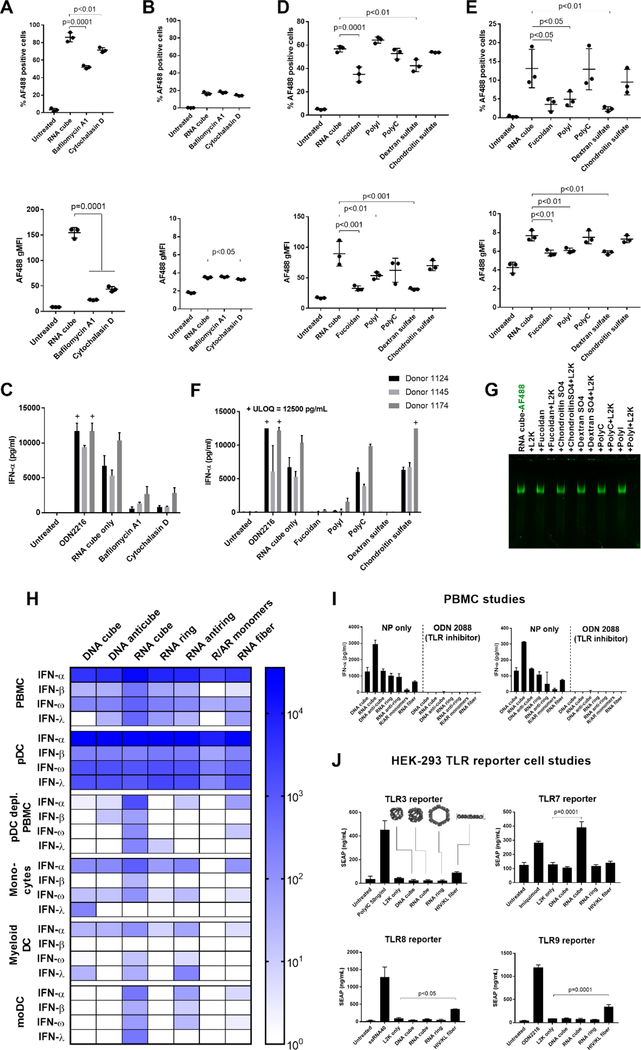
Nucleic acid nanoparticles (NANPs) have evolved as a new class of therapeutics with the potential to detect and treat diseases. Despite tremendous advancements in NANP development, their immunotoxicity, one of the major impediments in clinical translation of traditional therapeutic nucleic acids (TNAs), has never been fully characterized. Here, we describe the first systematically studied immunological recognition of 25 representative RNA and DNA NANPs selected to have different design principles and physicochemical properties. We discover that, unlike traditional TNAs, NANPs used without a delivery carrier are immunoquiescent. We show that interferons (IFNs) are the key cytokines triggered by NANPs after their internalization by phagocytic cells, which agrees with predictions based on the experiences with TNAs. However, in addition to type I IFNs, type III IFNs also serve as reliable biomarkers of NANPs, which is usually not characteristic of TNAs. We show that overall immunostimulation relies on NANP shapes, connectivities, and compositions. We demonstrate that, like with traditional TNAs, plasmacytoid dendritic cells serve as the primary interferon producers among all peripheral blood mononuclear cells treated with NANPs, and scavenger receptor-mediated uptake and endosomal Toll-like receptor signaling are essential for NANP immunorecognition. The TLR involvement, however, is different from that expected for traditional TNA recognition. Based on these results, we suggest that NANP technology may serve as a prototype of auxiliary molecular language for communication with the immune system and the modulation of immune responses.
Keywords: RNA and DNA nanoparticles; TLR; human PBMC; immunorecognition; interferons; pDC.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Toll-Like Receptor-Mediated Recognition of Nucleic Acid Nanoparticles (NANPs) in Human Primary Blood Cells.Molecules. 2019 Mar 20;24(6):1094. doi: 10.3390/molecules24061094. Molecules. 2019. PMID: 30897721 Free PMC article.
-
The Recognition of and Reactions to Nucleic Acid Nanoparticles by Human Immune Cells.Molecules. 2021 Jul 12;26(14):4231. doi: 10.3390/molecules26144231. Molecules. 2021. PMID: 34299506 Free PMC article. Review.
-
Change in Lipofectamine Carrier as a Tool to Fine-Tune Immunostimulation of Nucleic Acid Nanoparticles.Molecules. 2023 Jun 1;28(11):4484. doi: 10.3390/molecules28114484. Molecules. 2023. PMID: 37298960 Free PMC article.
-
Immunostimulation of Fibrous Nucleic Acid Nanoparticles Can be Modulated through Aptamer-Based Functional Moieties: Unveiling the Structure-Activity Relationship and Mechanistic Insights.ACS Appl Mater Interfaces. 2024 Feb 21;16(7):8430-8441. doi: 10.1021/acsami.3c17779. Epub 2024 Feb 12. ACS Appl Mater Interfaces. 2024. PMID: 38344840 Free PMC article.
-
Nucleic acid nanoparticles (NANPs) as molecular tools to direct desirable and avoid undesirable immunological effects.Adv Drug Deliv Rev. 2021 Jun;173:427-438. doi: 10.1016/j.addr.2021.04.011. Epub 2021 Apr 20. Adv Drug Deliv Rev. 2021. PMID: 33857556 Free PMC article. Review.
Cited by
-
Thermostability, Tunability, and Tenacity of RNA as Rubbery Anionic Polymeric Materials in Nanotechnology and Nanomedicine-Specific Cancer Targeting with Undetectable Toxicity.Chem Rev. 2021 Jul 14;121(13):7398-7467. doi: 10.1021/acs.chemrev.1c00009. Epub 2021 May 26. Chem Rev. 2021. PMID: 34038115 Free PMC article. Review.
-
Trends and patterns in cancer nanotechnology research: A survey of NCI's caNanoLab and nanotechnology characterization laboratory.Adv Drug Deliv Rev. 2022 Dec;191:114591. doi: 10.1016/j.addr.2022.114591. Epub 2022 Nov 1. Adv Drug Deliv Rev. 2022. PMID: 36332724 Free PMC article. Review.
-
RNA nanostructures for targeted drug delivery and imaging.RNA Biol. 2024 Jan;21(1):1-19. doi: 10.1080/15476286.2024.2328440. Epub 2024 Mar 31. RNA Biol. 2024. PMID: 38555519 Free PMC article. Review.
-
3D RNA-scaffolded wireframe origami.Nat Commun. 2023 Jan 24;14(1):382. doi: 10.1038/s41467-023-36156-1. Nat Commun. 2023. PMID: 36693871 Free PMC article.
-
Nanomaterials-Based Novel Immune Strategies in Clinical Translation for Cancer Therapy.Molecules. 2023 Jan 26;28(3):1216. doi: 10.3390/molecules28031216. Molecules. 2023. PMID: 36770883 Free PMC article. Review.
References
-
- Sasaki S; Guo S Nucleic Acid Ther. 2018, 28 (1), 1–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
