

Selective FcγR Co-engagement on APCs Modulates the Activity of Therapeutic Antibodies Targeting T Cell Antigens
- PMID: 29894690
- PMCID: PMC6292441
- DOI: 10.1016/j.ccell.2018.05.005
Selective FcγR Co-engagement on APCs Modulates the Activity of Therapeutic Antibodies Targeting T Cell Antigens
Abstract
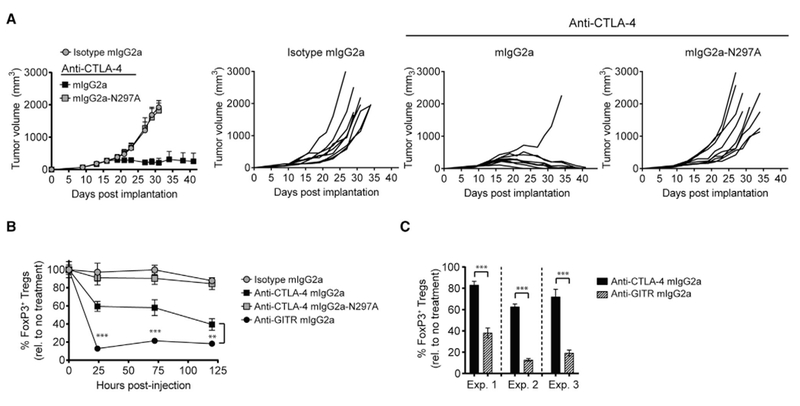
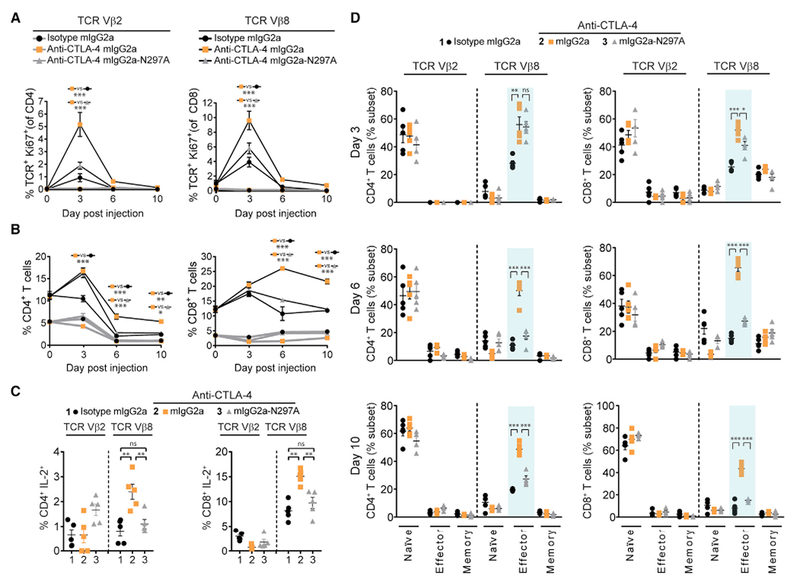
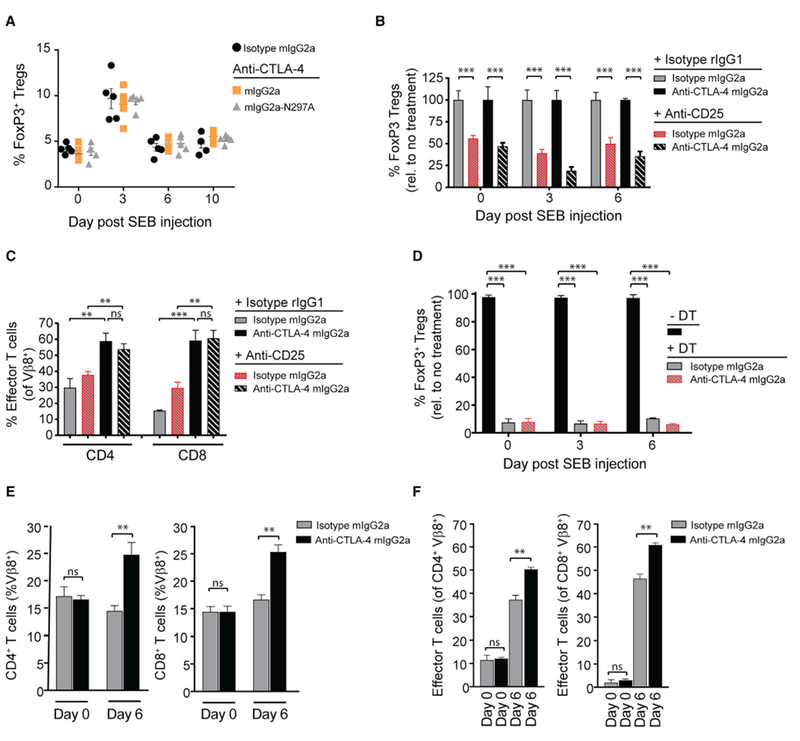
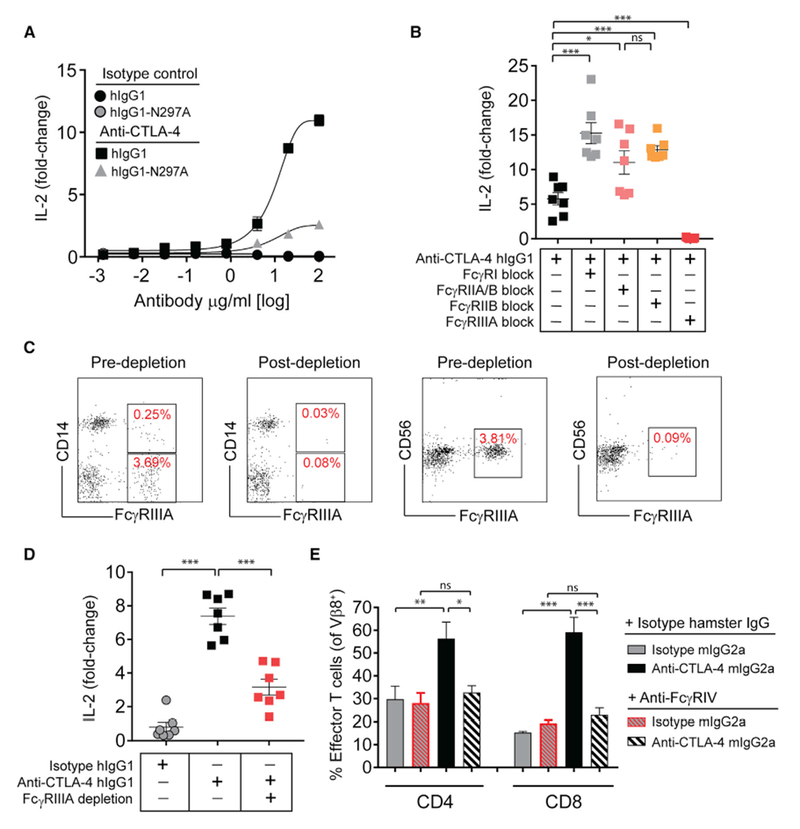
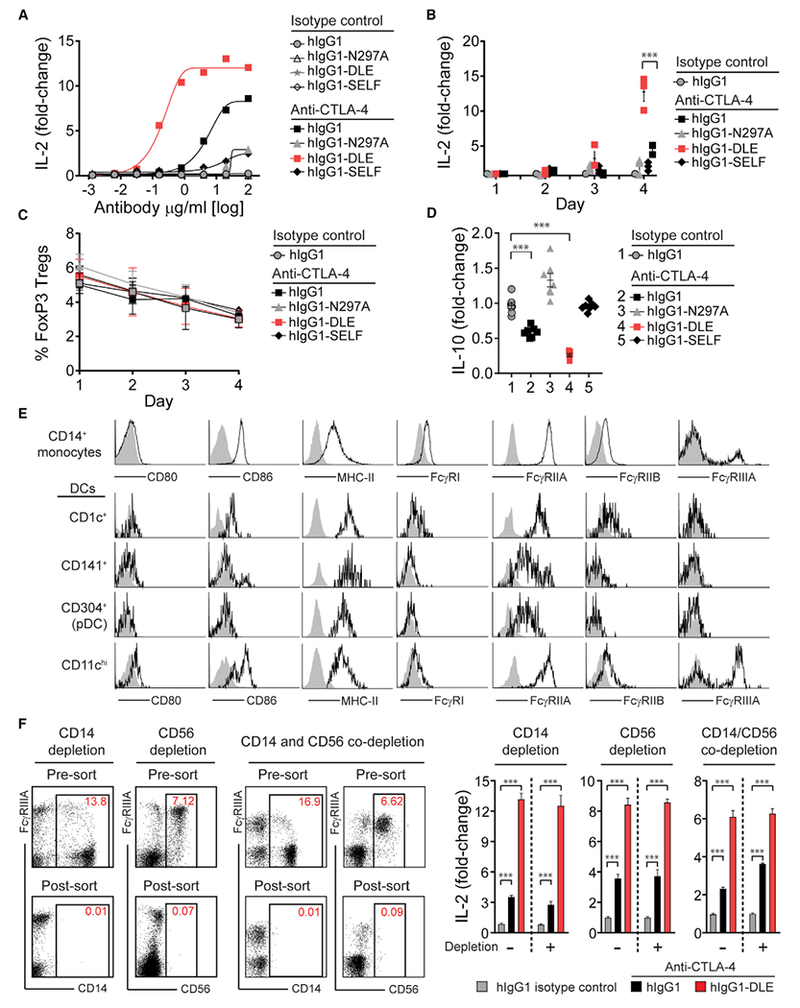
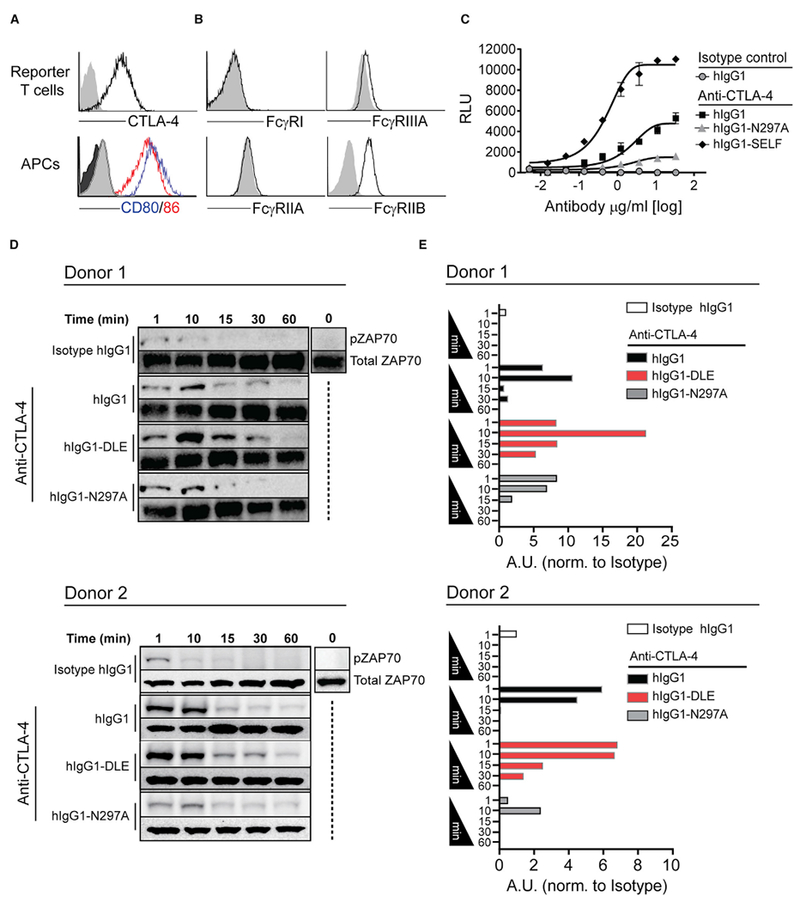
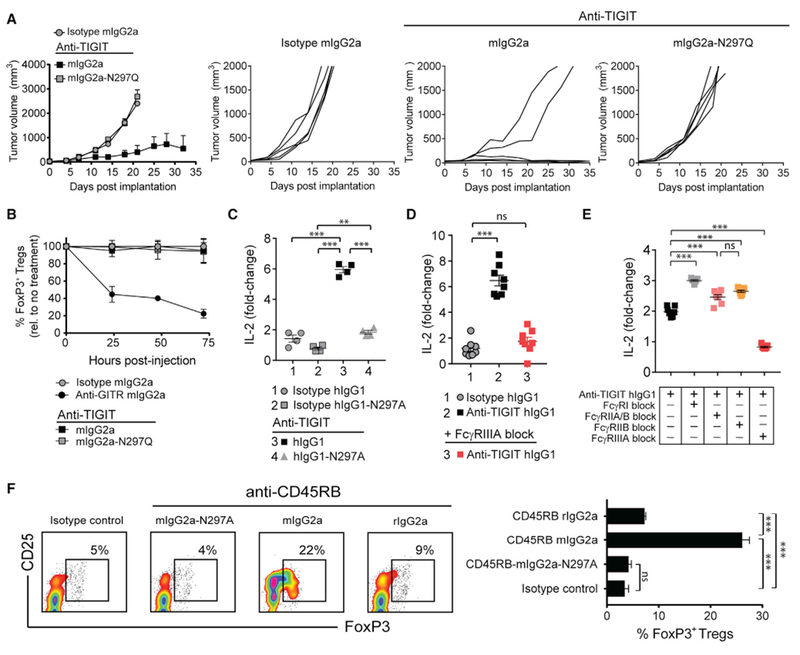
The co-engagement of fragment crystallizable (Fc) gamma receptors (FcγRs) with the Fc region of recombinant immunoglobulin monoclonal antibodies (mAbs) and its contribution to therapeutic activity has been extensively studied. For example, Fc-FcγR interactions have been shown to be important for mAb-directed effector cell activities, as well as mAb-dependent forward signaling into target cells via receptor clustering. Here we identify a function of mAbs targeting T cell-expressed antigens that involves FcγR co-engagement on antigen-presenting cells (APCs). In the case of mAbs targeting CTLA-4 and TIGIT, the interaction with FcγR on APCs enhanced antigen-specific T cell responses and tumoricidal activity. This mechanism extended to an anti-CD45RB mAb, which led to FcγR-dependent regulatory T cell expansion in mice.
Keywords: CD45RB; CTLA-4; Fc engineering; Fcγ receptor; TCR signaling; TIGIT; cancer immunotherapy; effector T cells; immune synapse; regulatory T cells.
Copyright © 2018 Elsevier Inc. All rights reserved.
Figures
References
-
- Acuto O, and Michel F (2003). CD28-mediated co-stimulation:aquantitative support for TCR signalling. Nat. Rev. Immunol 3, 939–951. - PubMed
-
- Arce Vargas F, Furness AJS, Solomon I, Joshi K, Mekkaoui L, Lesko MH, Miranda Rota E, Dahan R, Georgiou A, Sledzinska A, et al. (2017). Fc-optimized anti-CD25 depletes tumor-infiltrating regulatory T cells and synergizes with PD-1 blockade to eradicate established tumors. Immunity 46, 577–586. - PMC - PubMed
-
- Brownlie RJ, and Zamoyska R (2013). T cell receptor signalling networks: branched, diversified and bounded. Nat. Rev. Immunol 13, 257–269. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
