

The crystal structure of the Helicobacter pylori LlaJI.R1 N-terminal domain provides a model for site-specific DNA binding
- PMID: 29895618
- PMCID: PMC6066307
- DOI: 10.1074/jbc.RA118.001888
The crystal structure of the Helicobacter pylori LlaJI.R1 N-terminal domain provides a model for site-specific DNA binding
Abstract
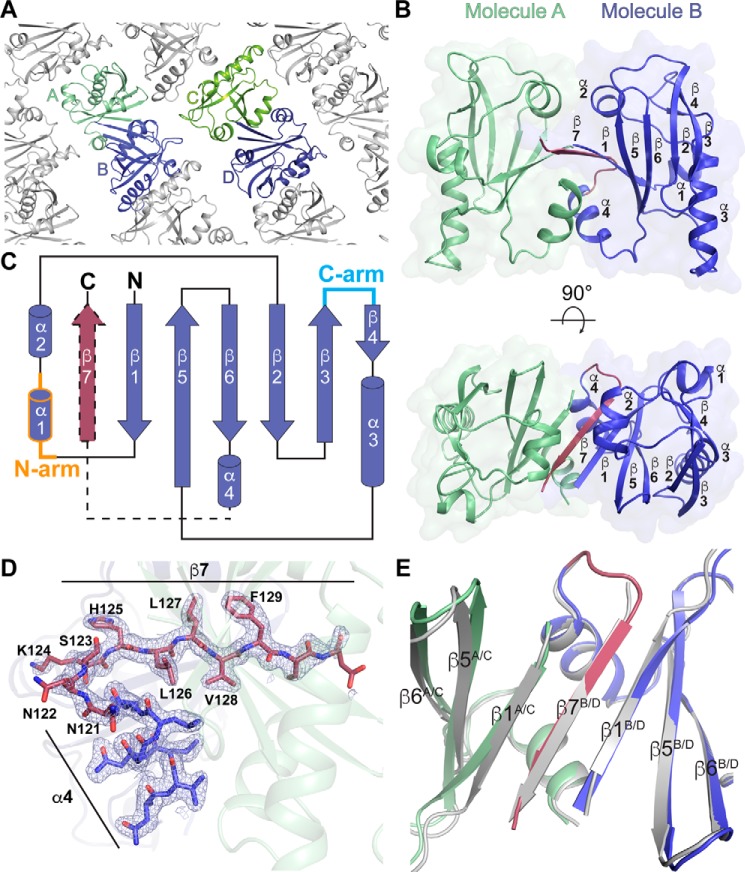
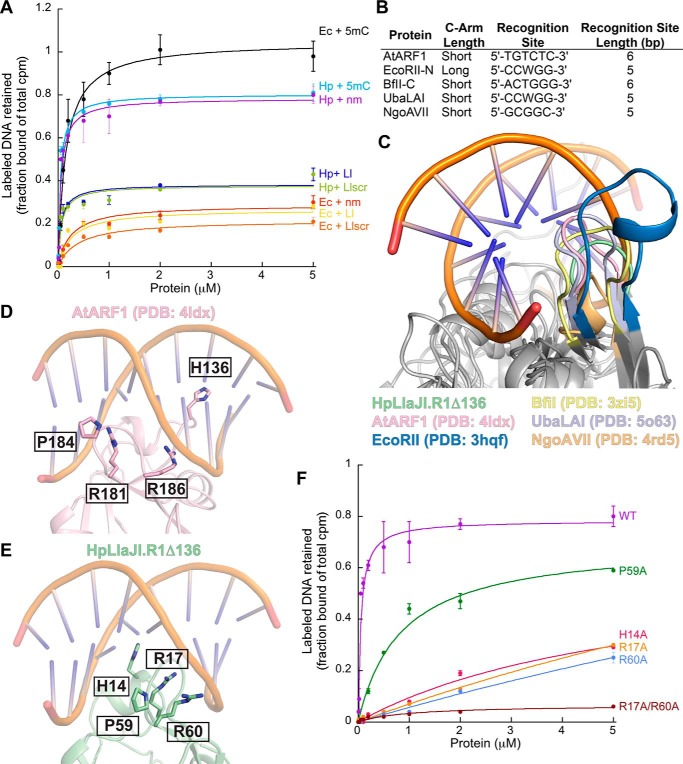
Restriction modification systems consist of an endonuclease that cleaves foreign DNA site-specifically and an associated methyltransferase that protects the corresponding target site in the host genome. Modification-dependent restriction systems, in contrast, specifically recognize and cleave methylated and/or glucosylated DNA. The LlaJI restriction system contains two 5-methylcytosine (5mC) methyltransferases (LlaJI.M1 and LlaJI.M2) and two restriction proteins (LlaJI.R1 and LlaJI.R2). LlaJI.R1 and LlaJI.R2 are homologs of McrB and McrC, respectively, which in Escherichia coli function together as a modification-dependent restriction complex specific for 5mC-containing DNA. Lactococcus lactis LlaJI.R1 binds DNA site-specifically, suggesting that the LlaJI system uses a different mode of substrate recognition. Here we present the structure of the N-terminal DNA-binding domain of Helicobacter pylori LlaJI.R1 at 1.97-Å resolution, which adopts a B3 domain fold. Structural comparison to B3 domains in plant transcription factors and other restriction enzymes identifies key recognition motifs responsible for site-specific DNA binding. Moreover, biochemistry and structural modeling provide a rationale for how H. pylori LlaJI.R1 may bind a target site that differs from the 5-bp sequence recognized by other LlaJI homologs and identify residues critical for this recognition activity. These findings underscore the inherent structural plasticity of B3 domains, allowing recognition of a variety of substrates using the same structural core.
Keywords: DNA binding protein; DNA endonuclease; DNA enzyme; X-ray crystallography; protein structure; restriction system.
© 2018 Hosford and Chappie.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures





Similar articles
-
The structure of the Thermococcus gammatolerans McrB N-terminal domain reveals a new mode of substrate recognition and specificity among McrB homologs.J Biol Chem. 2020 Jan 17;295(3):743-756. doi: 10.1074/jbc.RA119.010188. Epub 2019 Dec 10. J Biol Chem. 2020. PMID: 31822563 Free PMC article.
-
A genetic dissection of the LlaJI restriction cassette reveals insights on a novel bacteriophage resistance system.BMC Microbiol. 2006 Apr 28;6:40. doi: 10.1186/1471-2180-6-40. BMC Microbiol. 2006. PMID: 16646963 Free PMC article.
-
A dichotomous epigenetic mechanism governs expression of the LlaJI restriction/modification system.Mol Microbiol. 2005 Sep;57(6):1532-44. doi: 10.1111/j.1365-2958.2005.04769.x. Mol Microbiol. 2005. PMID: 16135222
-
Restriction-modification systems may be associated with Helicobacter pylori virulence.J Gastroenterol Hepatol. 2010 May;25 Suppl 1:S95-8. doi: 10.1111/j.1440-1746.2009.06211.x. J Gastroenterol Hepatol. 2010. PMID: 20586875 Review.
-
Sticky socks: Helicobacter pylori VacA takes shape.Trends Microbiol. 2008 Mar;16(3):89-92. doi: 10.1016/j.tim.2008.01.001. Epub 2008 Feb 14. Trends Microbiol. 2008. PMID: 18280164 Review.
Cited by
-
The structure of the Thermococcus gammatolerans McrB N-terminal domain reveals a new mode of substrate recognition and specificity among McrB homologs.J Biol Chem. 2020 Jan 17;295(3):743-756. doi: 10.1074/jbc.RA119.010188. Epub 2019 Dec 10. J Biol Chem. 2020. PMID: 31822563 Free PMC article.
-
Rapid Detection of Helicobacter pylori by the Naked Eye Using DNA Aptamers.ACS Omega. 2021 Jan 21;6(5):3771-3779. doi: 10.1021/acsomega.0c05374. eCollection 2021 Feb 9. ACS Omega. 2021. PMID: 33585756 Free PMC article.
-
Structural asymmetry governs the assembly and GTPase activity of McrBC restriction complexes.Nat Commun. 2020 Nov 20;11(1):5907. doi: 10.1038/s41467-020-19735-4. Nat Commun. 2020. PMID: 33219217 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
