

Genetic Determinants Associated With in Vivo Survival of Burkholderia cenocepacia in the Caenorhabditis elegans Model
- PMID: 29896180
- PMCID: PMC5987112
- DOI: 10.3389/fmicb.2018.01118
Genetic Determinants Associated With in Vivo Survival of Burkholderia cenocepacia in the Caenorhabditis elegans Model
Abstract
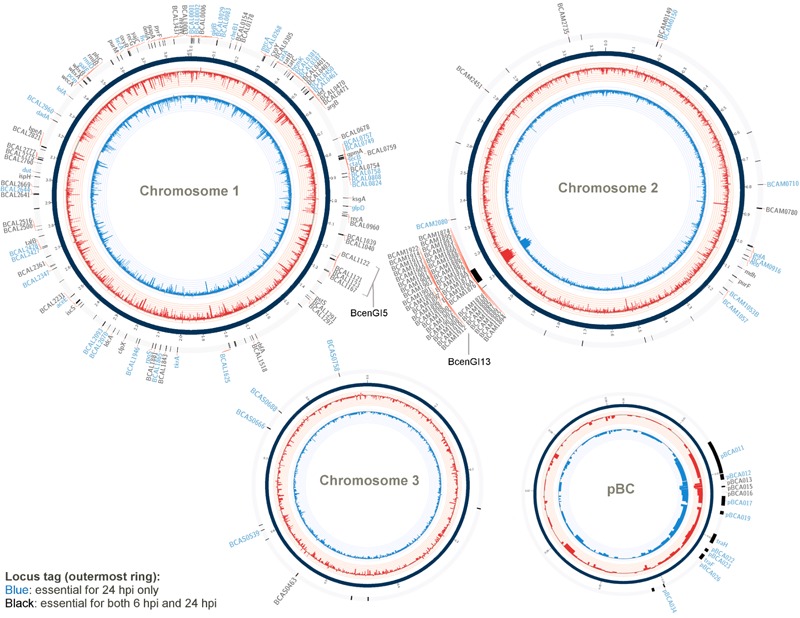
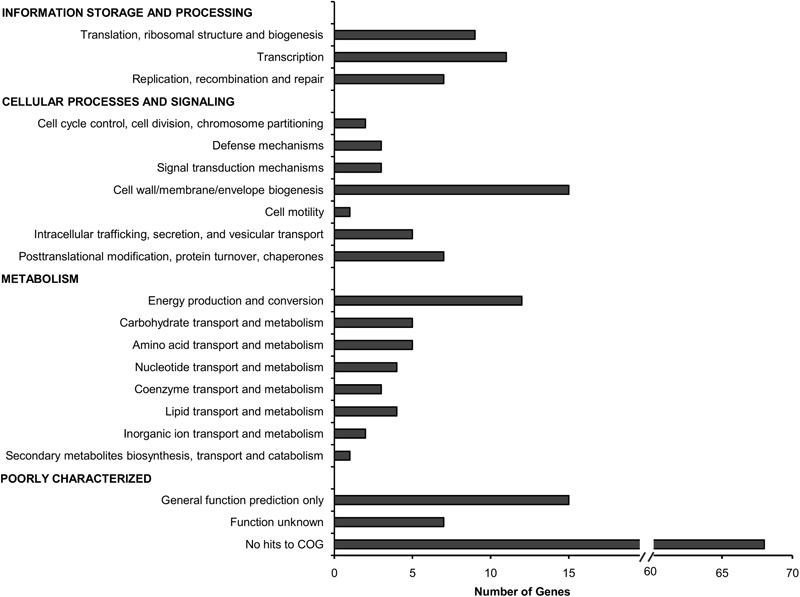
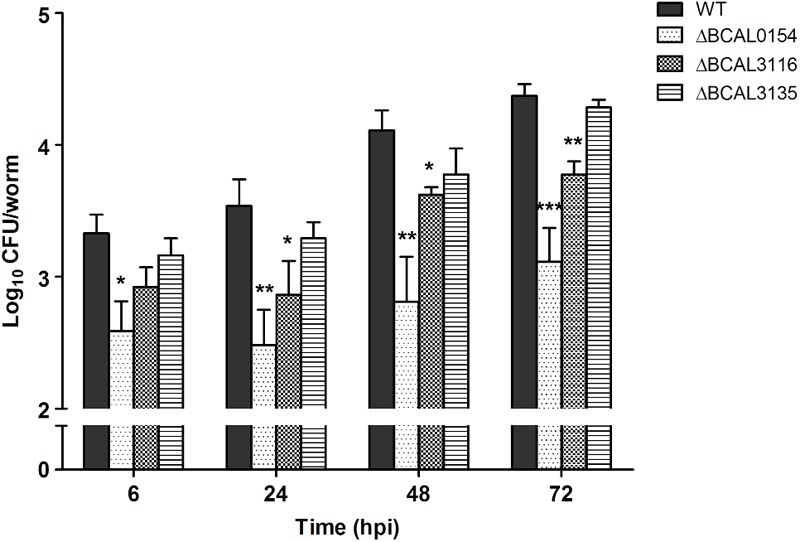
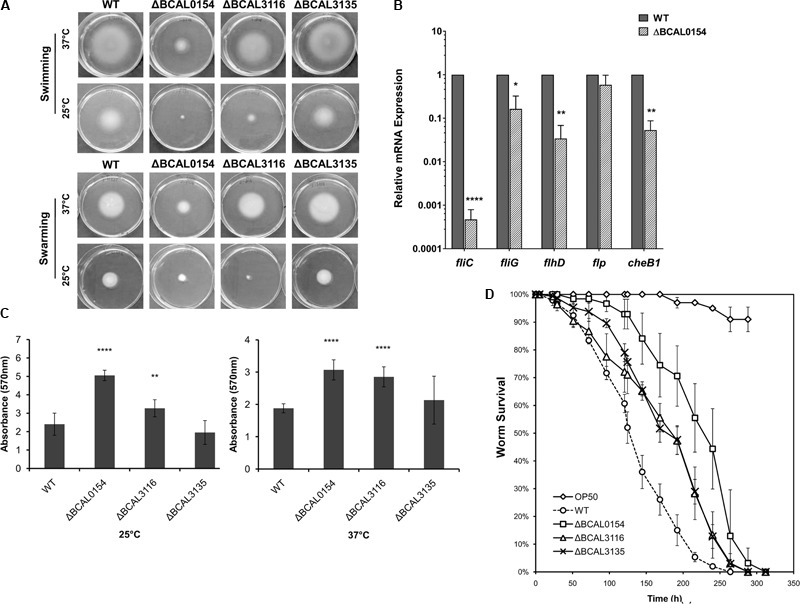
A Burkholderia cenocepacia infection usually leads to reduced survival and fatal cepacia syndrome in cystic fibrosis patients. The identification of B. cenocepacia essential genes for in vivo survival is key to designing new anti-infectives therapies. We used the Transposon-Directed Insertion Sequencing (TraDIS) approach to identify genes required for B. cenocepacia survival in the model infection host, Caenorhabditis elegans. A B. cenocepacia J2315 transposon pool of ∼500,000 mutants was used to infect C. elegans. We identified 178 genes as crucial for B. cenocepacia survival in the infected nematode. The majority of these genes code for proteins of unknown function, many of which are encoded by the genomic island BcenGI13, while other gene products are involved in nutrient acquisition, general stress responses and LPS O-antigen biosynthesis. Deletion of the glycosyltransferase gene wbxB and a histone-like nucleoid structuring (H-NS) protein-encoding gene (BCAL0154) reduced bacterial accumulation and attenuated virulence in C. elegans. Further analysis using quantitative RT-PCR indicated that BCAL0154 modulates B. cenocepacia pathogenesis via transcriptional regulation of motility-associated genes including fliC, fliG, flhD, and cheB1. This screen has successfully identified genes required for B. cenocepacia survival within the host-associated environment, many of which are potential targets for developing new antimicrobials.
Keywords: Burkholderia cenocepacia; Caenorhabditis elegans; TraDIS; essential genes; in vivo survival.
Figures
Similar articles
-
Identification of Burkholderia cenocepacia non-coding RNAs expressed during Caenorhabditis elegans infection.Appl Microbiol Biotechnol. 2023 Jun;107(11):3653-3671. doi: 10.1007/s00253-023-12530-3. Epub 2023 Apr 25. Appl Microbiol Biotechnol. 2023. PMID: 37097504 Free PMC article.
-
Candidate Essential Genes in Burkholderia cenocepacia J2315 Identified by Genome-Wide TraDIS.Front Microbiol. 2016 Aug 22;7:1288. doi: 10.3389/fmicb.2016.01288. eCollection 2016. Front Microbiol. 2016. PMID: 27597847 Free PMC article.
-
Unraveling Burkholderia cenocepacia H111 fitness determinants using two animal models.mSystems. 2025 Apr 22;10(4):e0135424. doi: 10.1128/msystems.01354-24. Epub 2025 Mar 19. mSystems. 2025. PMID: 40105327 Free PMC article.
-
Burkholderia cenocepacia in cystic fibrosis: epidemiology and molecular mechanisms of virulence.Clin Microbiol Infect. 2010 Jul;16(7):821-30. doi: 10.1111/j.1469-0691.2010.03237.x. Clin Microbiol Infect. 2010. PMID: 20880411 Review.
-
Molecular approaches to pathogenesis study of Burkholderia cenocepacia, an important cystic fibrosis opportunistic bacterium.Appl Microbiol Biotechnol. 2011 Dec;92(5):887-95. doi: 10.1007/s00253-011-3616-5. Epub 2011 Oct 14. Appl Microbiol Biotechnol. 2011. PMID: 21997606 Review.
Cited by
-
Identification of Burkholderia cenocepacia non-coding RNAs expressed during Caenorhabditis elegans infection.Appl Microbiol Biotechnol. 2023 Jun;107(11):3653-3671. doi: 10.1007/s00253-023-12530-3. Epub 2023 Apr 25. Appl Microbiol Biotechnol. 2023. PMID: 37097504 Free PMC article.
-
Methodological tools to study species of the genus Burkholderia.Appl Microbiol Biotechnol. 2021 Dec;105(24):9019-9034. doi: 10.1007/s00253-021-11667-3. Epub 2021 Nov 10. Appl Microbiol Biotechnol. 2021. PMID: 34755214 Free PMC article. Review.
-
Phenylacetyl Coenzyme A, Not Phenylacetic Acid, Attenuates CepIR-Regulated Virulence in Burkholderia cenocepacia.Appl Environ Microbiol. 2019 Nov 27;85(24):e01594-19. doi: 10.1128/AEM.01594-19. Print 2019 Dec 15. Appl Environ Microbiol. 2019. PMID: 31585996 Free PMC article.
-
A Histone-Like Nucleoid Structuring Protein Regulates Several Virulence Traits in Burkholderia multivorans.Appl Environ Microbiol. 2021 Jun 25;87(14):e0036921. doi: 10.1128/AEM.00369-21. Epub 2021 Jun 25. Appl Environ Microbiol. 2021. PMID: 33931418 Free PMC article.
-
Genome-wide transposon mutagenesis analysis of Burkholderia pseudomallei reveals essential genes for in vitro and in vivo survival.Front Cell Infect Microbiol. 2022 Dec 23;12:1062682. doi: 10.3389/fcimb.2022.1062682. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 36619746 Free PMC article.
References
-
- Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 57 289–300.
-
- Browning D. F., Wells T. J., França F. L. S., Morris F. C., Sevastsyanovich Y. R., Bryant J. A., et al. (2013). Laboratory adapted Escherichia coli K-12 becomes a pathogen of Caenorhabditis elegans upon restoration of O antigen biosynthesis. Mol. Microbiol. 87 939–950. 10.1111/mmi.12144 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous