

The Lectin Complement Pathway Is Involved in Protection Against Enteroaggregative Escherichia coli Infection
- PMID: 29896194
- PMCID: PMC5986924
- DOI: 10.3389/fimmu.2018.01153
The Lectin Complement Pathway Is Involved in Protection Against Enteroaggregative Escherichia coli Infection
Abstract
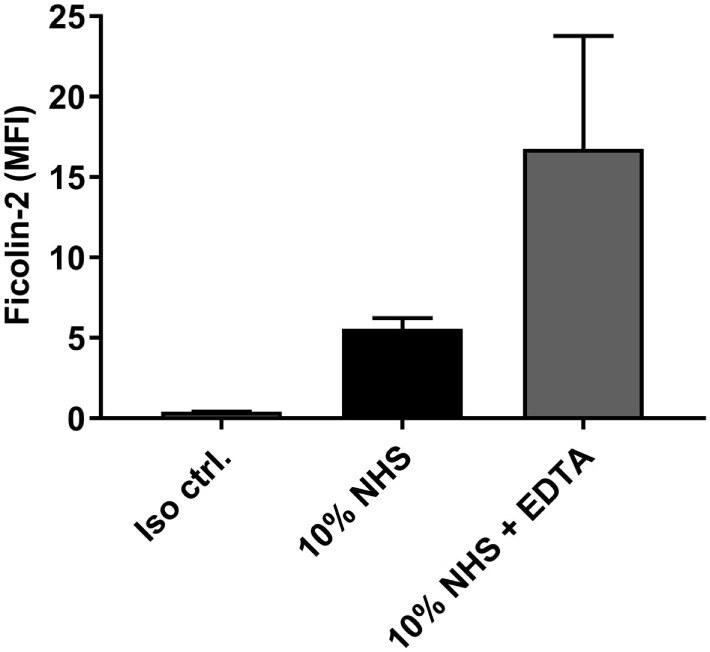
Enteroaggregative Escherichia coli (EAEC) causes acute and persistent diarrhea worldwide. Still, the involvement of host factors in EAEC infections is unresolved. Binding of recognition molecules from the lectin pathway of complement to EAEC strains have been observed, but the importance is not known. Our aim was to uncover the involvement of these molecules in innate complement dependent immune protection toward EAEC. Binding of mannose-binding lectin, ficolin-1, -2, and -3 to four prototypic EAEC strains, and ficolin-2 binding to 56 clinical EAEC isolates were screened by a consumption-based ELISA method. Flow cytometry was used to determine deposition of C4b, C3b, and the bactericidal C5b-9 membrane attack complex (MAC) on the bacteria in combination with different complement inhibitors. In addition, the direct serum bactericidal effect was assessed. Screening of the prototypic EAEC strains revealed that ficolin-2 was the major binder among the lectin pathway recognition molecules. However, among the clinical EAEC isolates only a restricted number (n = 5) of the isolates bound ficolin-2. Using the ficolin-2 binding isolate C322-17 as a model, we found that incubation with normal human serum led to deposition of C4b, C3b, and to MAC formation. No inhibition of complement deposition was observed when a C1q inhibitor was added, while partial inhibition was observed when ficolin-2 or factor D inhibitors were used separately. Combining the inhibitors against ficolin-2 and factor D led to virtually complete inhibition of complement deposition and protection against direct bacterial killing. These results demonstrate that ficolin-2 may play an important role in innate immune protection against EAEC when an appropriate ligand is exposed, but many EAEC strains evade lectin pathway recognition and may, therefore, circumvent this strategy of innate host immune protection.
Keywords: enteroaggregative Escherichia coli, complement; ficolin-2; lectin pathway; serum resistance.
Figures








Similar articles
-
Studies of the binding of ficolin-2 and ficolin-3 from the complement lectin pathway to Leptospira biflexa, Pasteurella pneumotropica and Diarrheagenic Escherichia coli.Immunobiology. 2015 Oct;220(10):1177-85. doi: 10.1016/j.imbio.2015.06.001. Epub 2015 Jun 4. Immunobiology. 2015. PMID: 26074063
-
A novel measurement method for activation of the lectin complement pathway via both mannose-binding lectin (MBL) and L-ficolin.J Immunol Methods. 2009 Sep 30;349(1-2):9-17. doi: 10.1016/j.jim.2009.08.005. Epub 2009 Aug 20. J Immunol Methods. 2009. PMID: 19699205
-
Cholesterol Crystals Activate the Lectin Complement Pathway via Ficolin-2 and Mannose-Binding Lectin: Implications for the Progression of Atherosclerosis.J Immunol. 2016 Jun 15;196(12):5064-74. doi: 10.4049/jimmunol.1502595. Epub 2016 May 16. J Immunol. 2016. PMID: 27183610
-
A journey through the lectin pathway of complement-MBL and beyond.Immunol Rev. 2016 Nov;274(1):74-97. doi: 10.1111/imr.12468. Immunol Rev. 2016. PMID: 27782323 Review.
-
MBL2, FCN1, FCN2 and FCN3-The genes behind the initiation of the lectin pathway of complement.Mol Immunol. 2009 Sep;46(14):2737-44. doi: 10.1016/j.molimm.2009.05.005. Epub 2009 Jun 7. Mol Immunol. 2009. PMID: 19501910 Review.
Cited by
-
Decreased Ficolin-3-mediated Complement Lectin Pathway Activation and Alternative Pathway Amplification During Bacterial Infections in Patients With Type 2 Diabetes Mellitus.Front Immunol. 2019 Mar 20;10:509. doi: 10.3389/fimmu.2019.00509. eCollection 2019. Front Immunol. 2019. PMID: 30949171 Free PMC article.
-
Antibodies Specific to Membrane Proteins Are Effective in Complement-Mediated Killing of Mycoplasma bovis.Infect Immun. 2019 Nov 18;87(12):e00740-19. doi: 10.1128/IAI.00740-19. Print 2019 Dec. Infect Immun. 2019. PMID: 31548318 Free PMC article.
-
C9 immunostaining as a tissue biomarker for periprosthetic joint infection diagnosis.Front Immunol. 2023 Feb 21;14:1112188. doi: 10.3389/fimmu.2023.1112188. eCollection 2023. Front Immunol. 2023. PMID: 36895567 Free PMC article.
-
Biochemical and structural characterization of a recombinant fibrinogen-related lectin from Penaeus monodon.Sci Rep. 2021 Feb 3;11(1):2934. doi: 10.1038/s41598-021-82301-5. Sci Rep. 2021. PMID: 33536457 Free PMC article.
-
Ficolins and the Recognition of Pathogenic Microorganisms: An Overview of the Innate Immune Response and Contribution of Single Nucleotide Polymorphisms.J Immunol Res. 2019 Feb 5;2019:3205072. doi: 10.1155/2019/3205072. eCollection 2019. J Immunol Res. 2019. PMID: 30868077 Free PMC article. Review.
References
-
- Hebbelstrup Jensen B, Poulsen A, Hebbelstrup Rye Rasmussen S, Struve C, Engberg JH, Friis-Møller A, et al. Genetic virulence profile of enteroaggregative Escherichia coli strains isolated from Danish children with either acute or persistent diarrhea. Front Cell Infect Microbiol (2017) 7:230.10.3389/fcimb.2017.00230 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous