

Review
doi: 10.1152/physiol.00014.2018.
Carotid Bodies and the Integrated Cardiorespiratory Response to Hypoxia
Affiliations
- PMID: 29897299
- PMCID: PMC6088141
- DOI: 10.1152/physiol.00014.2018
Item in Clipboard
Review
Carotid Bodies and the Integrated Cardiorespiratory Response to Hypoxia
Physiology (Bethesda).
.
Abstract
Advances in our understanding of brain mechanisms for the hypoxic ventilatory response, coordinated changes in blood pressure, and the long-term consequences of chronic intermittent hypoxia as in sleep apnea, such as hypertension and heart failure, are giving impetus to the search for therapies to "erase" dysfunctional memories distributed in the carotid bodies and central nervous system. We review current network models, open questions, sex differences, and implications for translational research.
Figures
Continued
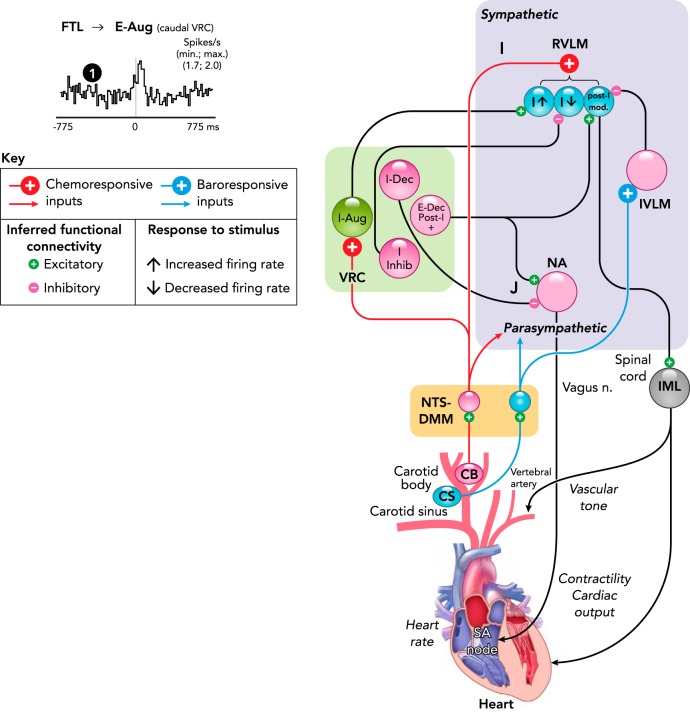
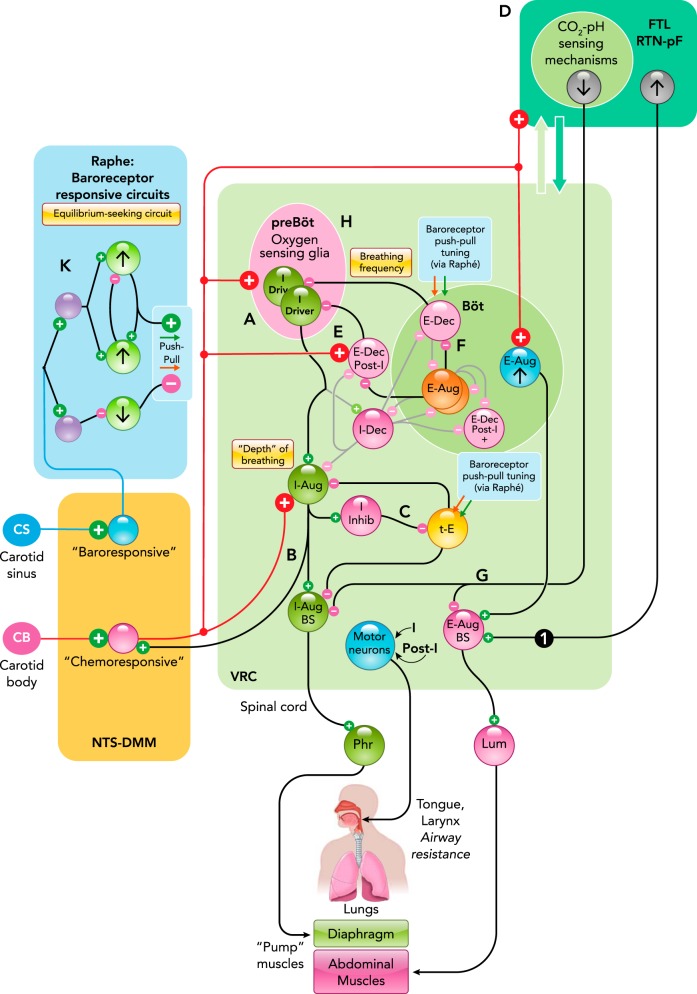
Medullary circuit mechanisms in contemporary models of carotid chemoreceptor and baroreceptor modulation of breathing and cardiovascular functions A–K: schematic representations of specific functional interactions described in the text. Neuron populations are grouped by brain stem location and labeled by their respiratory modulated firing rates: inspiratory (I), post-inspiratory (post-I), or expiratory (E), according to the phase of greatest average activity, and as decrementing (Dec) or augmenting (Aug) if the average peak firing rate occurs during the first or second half of the phase, respectively. I-Driver neuron activity begins slightly before phrenic nerve discharge, with firing rates that peak in the early I phase and then slowly decrease before abruptly decreasing at the end of inspiration. Tonic E cells (t-E) are active throughout the respiratory cycle, with their greatest rate during expiration. Black lines and “synapses” indicate inferred functional connectivity discussed in the text. 1, Cross-correlogram with offset peak consistent with RTN-pF region neuron exciting a putative premotor caudal VRC augmenting expiratory neuron. Figure adapted from Fig. 5D in Ref. with permission from Journal of Neurophysiology. Both neurons responded to transient hypoxia with increased firing rates (177). Binwidth = 15.5 ms, 71,132 trigger neuron spikes, 25,541 target neuron spikes. Böt, Bötzinger complex; BS, bulbospinal; FTL/RTN-pF, lateral tegmental field/retrotrapezoid n.-parafacial n; IML, intermediolateral column of the spinal cord; IVLM, intermediate ventrolateral medulla; Lum, lumbar nerve; NA, n. ambiguus; NTS-DMM, n. tractus solitarius-dorsal medial medulla; Phr, phrenic nerve; pre-Böt, pre-Bötzinger complex; RVLM, rostral ventrolateral medulla; VRC, ventral respiratory column.
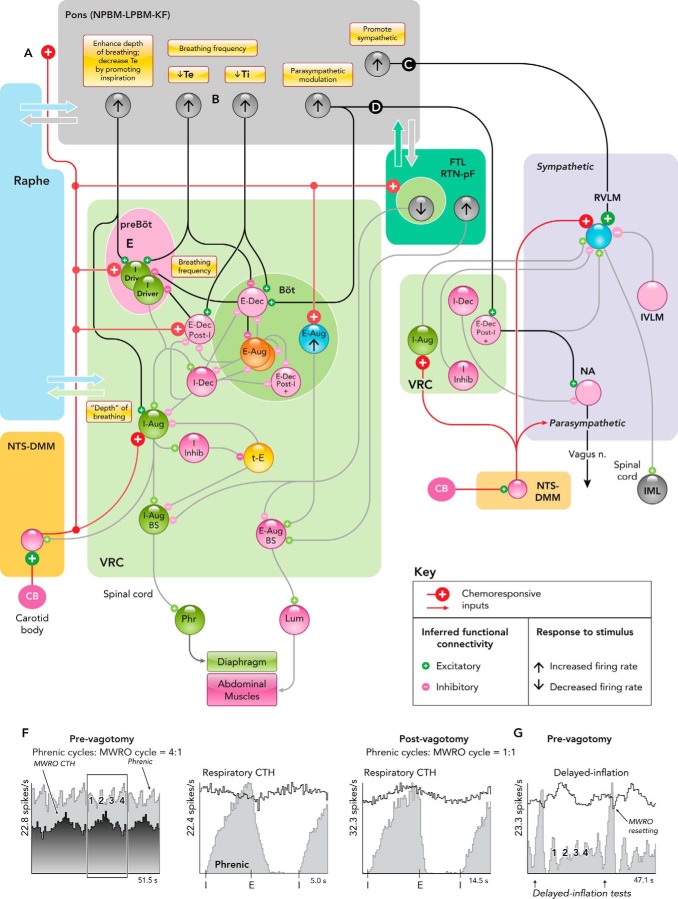
Simplified functional connectivity of hypoxia-responsive pontine neurons A–E: interactions inferred from various experimental approaches reviewed in the text that contribute to the hypoxic ventilatory response and changes in cardiorespiratory coupling during hypoxia in current models. F and G: MWRO-triggered firing rate histograms of a pontine neuron before and after bilateral vagotomy and phase resetting of MWROs by withholding lung inflation in vagus-intact cat. Figure derived from Fig. 5A in Ref. with permission. See text for details. KF, Kölliker-Fuse n.; LPBM, lateral parabrachial n.; MWRO, Mayer wave-related oscillations; NPBM, medial parabrachial n.; Ti (Te), duration of inspiratory (expiratory) phase.
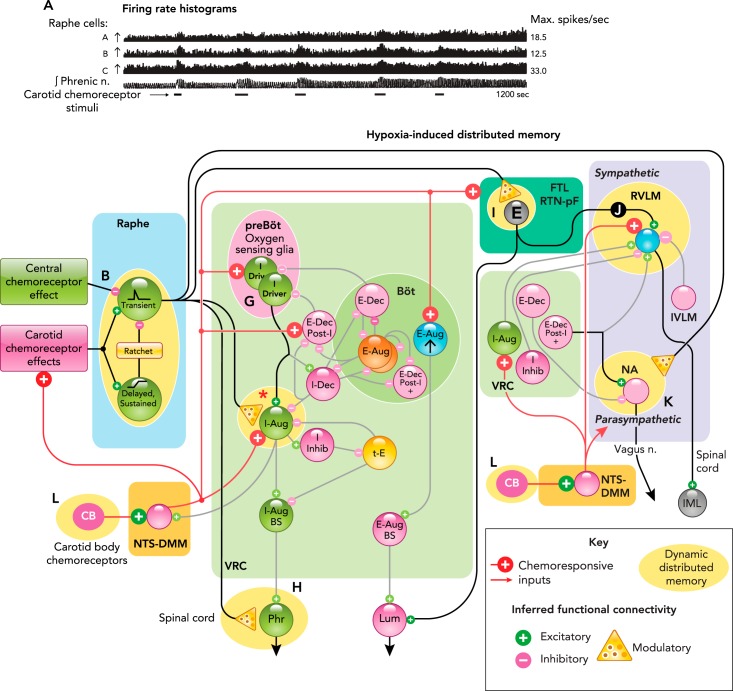
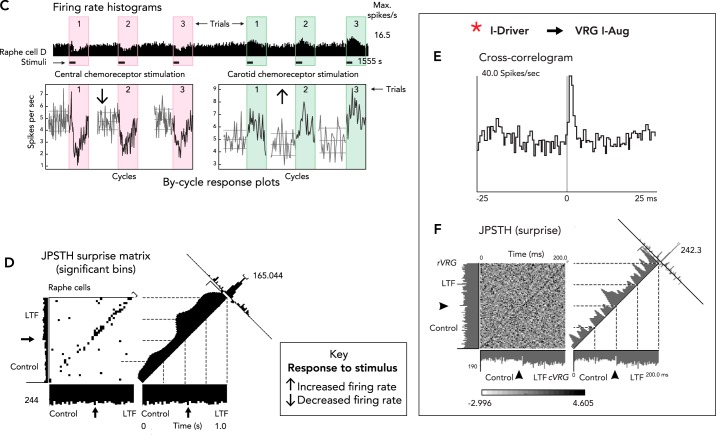
Distributed network sites for induction and expression of carotid body- and intermittent hypoxia-induced long-term facilitation A: responses of raphe neurons and integrated phrenic nerve activity during repeated selective stimulation of carotid chemoreceptors and induction of long-term facilitation (LTF). Figure is derived from Ref. with permission from Respiration Physiology. B: raphe circuit mechanism proposed to contribute to ratchet-like incremental induction of LTF (171, 172). C: firing rate histogram and by-cycle rate plots show opposite responses of a raphe neuron to selective stimulation of central and peripheral carotid chemoreceptors. Figure derived from Fig. 2A, C in Ref. with permission from The Royal Society. D: increased number of black bins in the top right quadrant along the diagonal of the joint peri-stimulus time histogram (JPSTH) documents a significant increase in spike synchrony and effective connectivity for two raphe cells following induction of LTF. Figure adapted from Fig. 4A in Ref. with permission from Journal of Physiology. E: offset cross-correlogram peak shows effective connectivity between a pre-Bötzinger I-Driver neuron and a VRG inspiratory target cell; binwidth = 0.5 ms, 8,216 trigger neuron spikes, 4,569 target neuron spikes. F: non-uniform distribution of statistically significant black bins along the diagonal of the JPSTH for the same neurons shown in E documents enhanced effective connectivity between this pair of neurons following induction of LTF. Figure derived from Fig. 3, B and C, in Ref. with permission from Journal of Physiology. G–L: additional sites for distributed LTF expression following repeated episodes of intermittent hypoxia. See text for details.
Continued
References
-
- Arata A, Tanaka I, Fujii M, Ezure K. Active inspiratory-expiratory phase switching mechanism exists in the neonatal nucleus parabrachialis. In: New Frontiers in Respiratory Control. Advances in Experimental Medicine and Biology, edited by Homma I, Onimaru H, Fukuchi Y. New York, NY: Springer New York, 2010, p. 135–138. doi: 10.1007/978-1-4419-5692-7_27. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources