

Understanding the Agility of Running Birds: Sensorimotor and Mechanical Factors in Avian Bipedal Locomotion
- PMID: 29897448
- PMCID: PMC6204994
- DOI: 10.1093/icb/icy058
Understanding the Agility of Running Birds: Sensorimotor and Mechanical Factors in Avian Bipedal Locomotion
Abstract
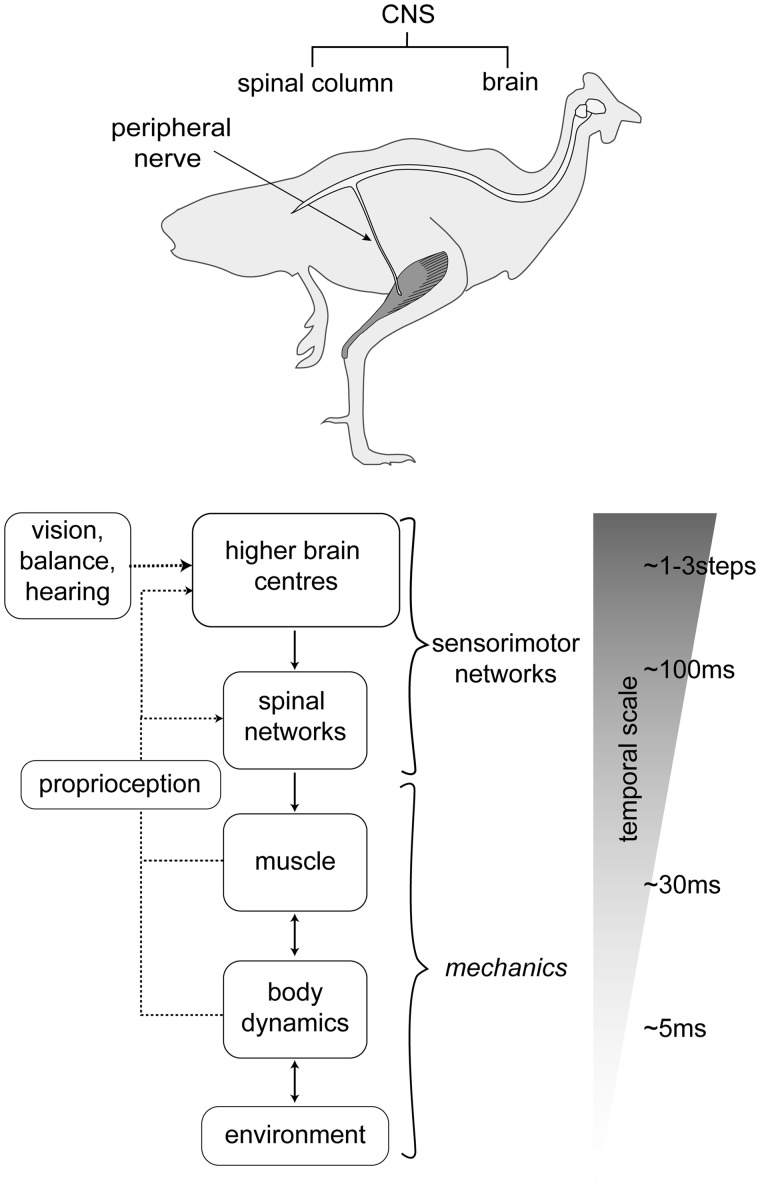
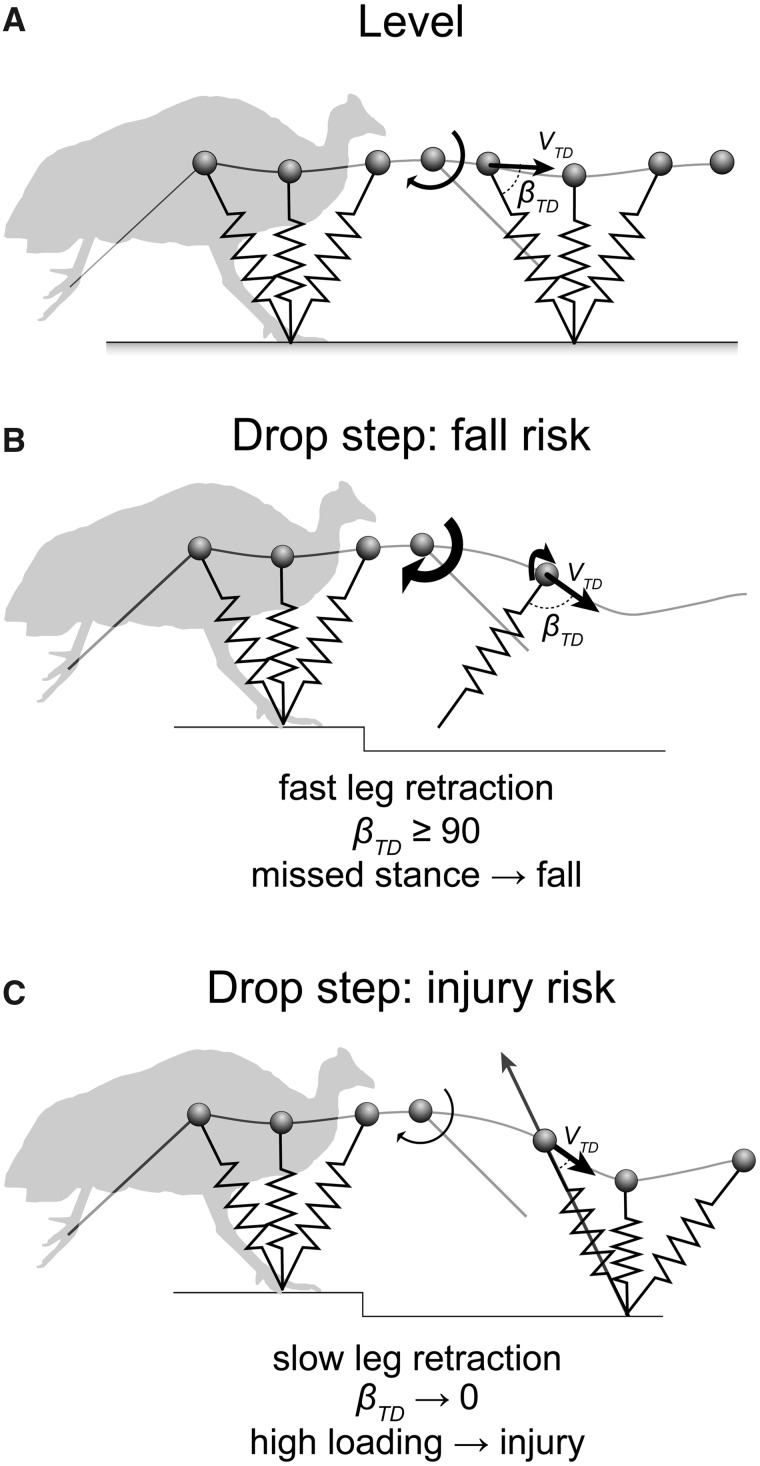
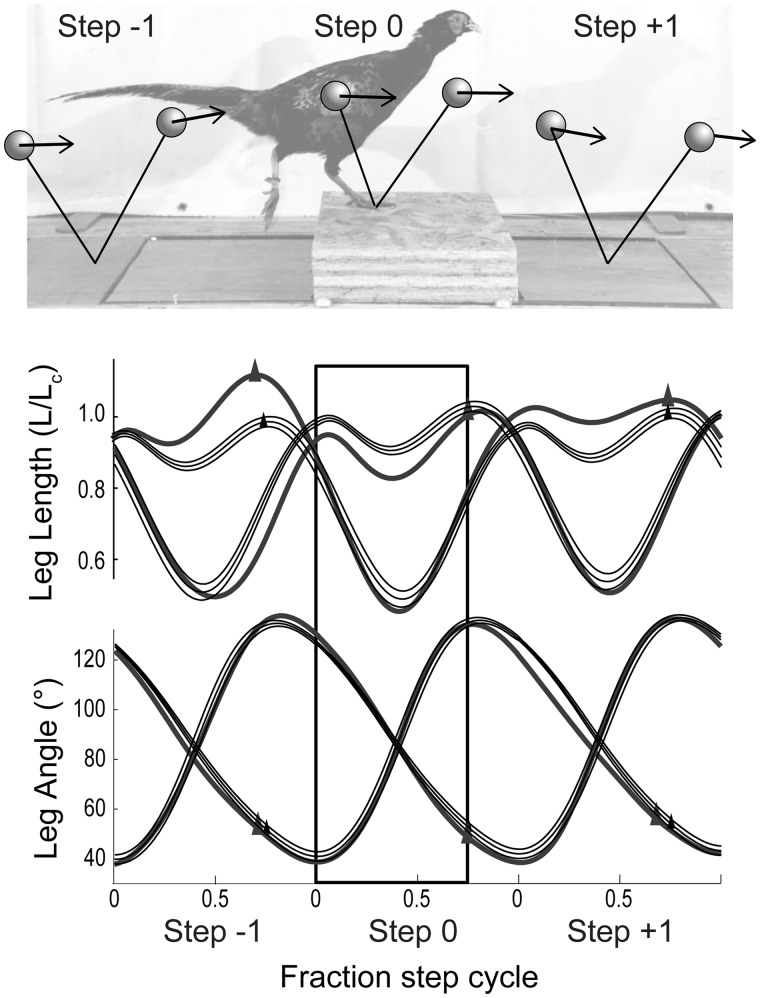
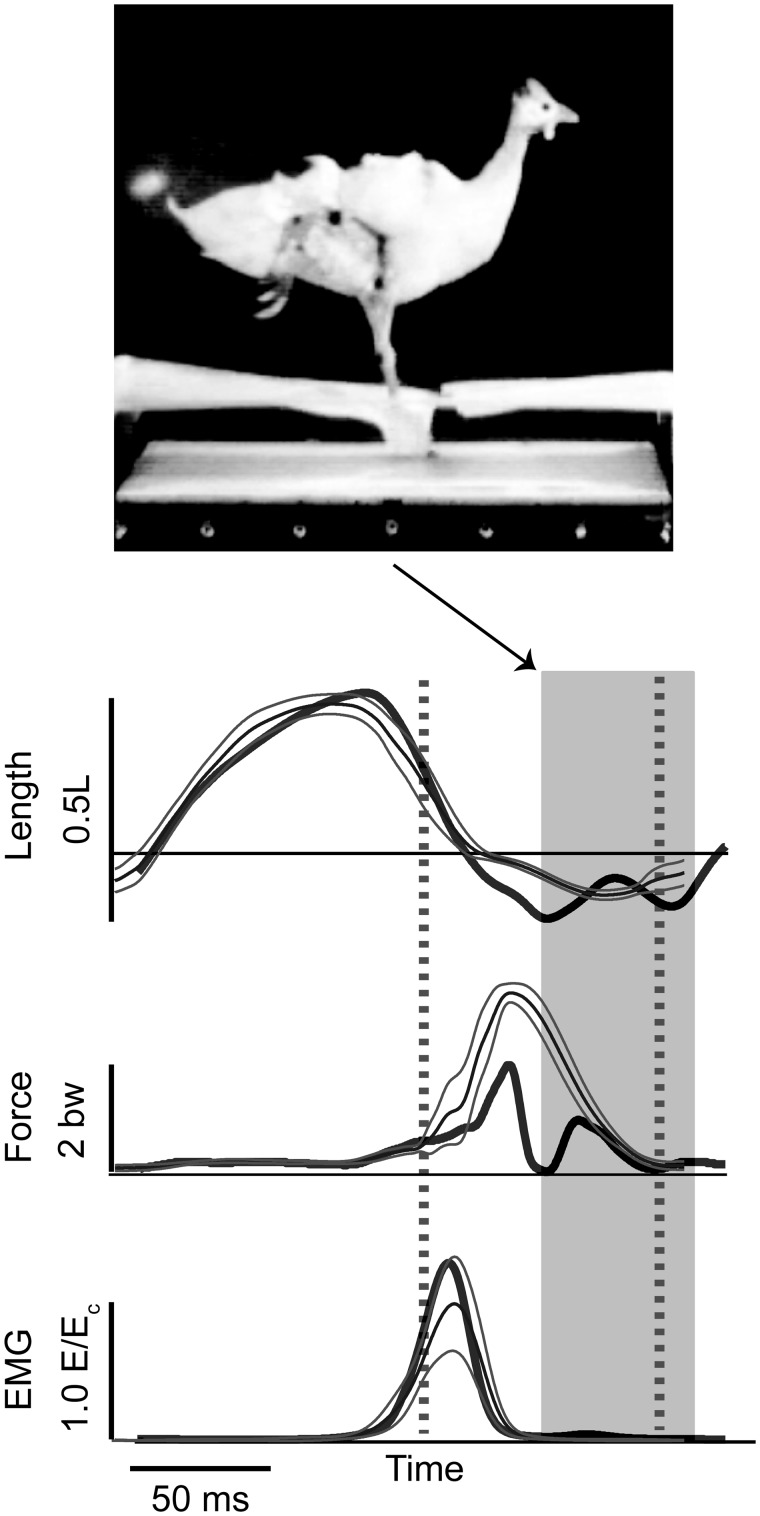
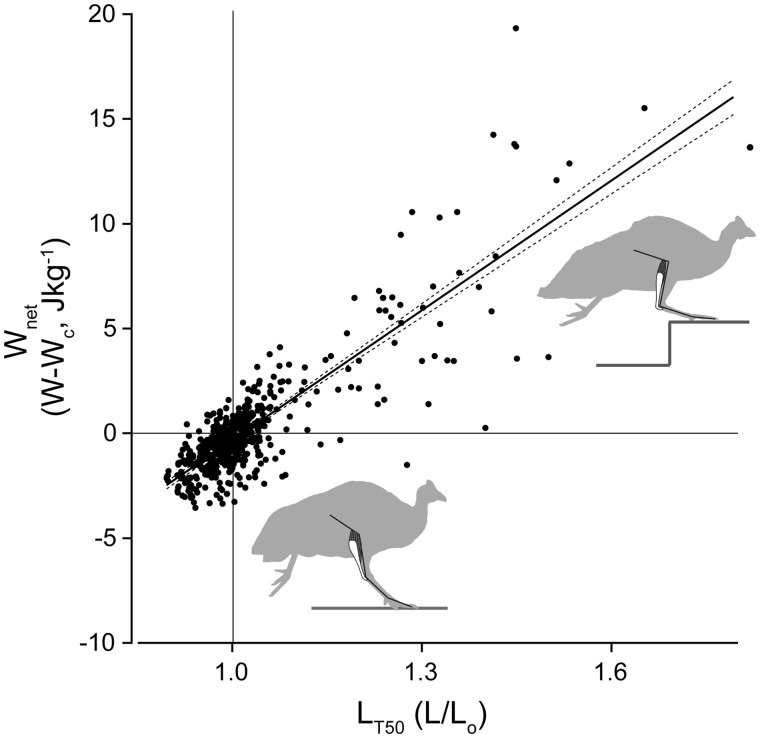
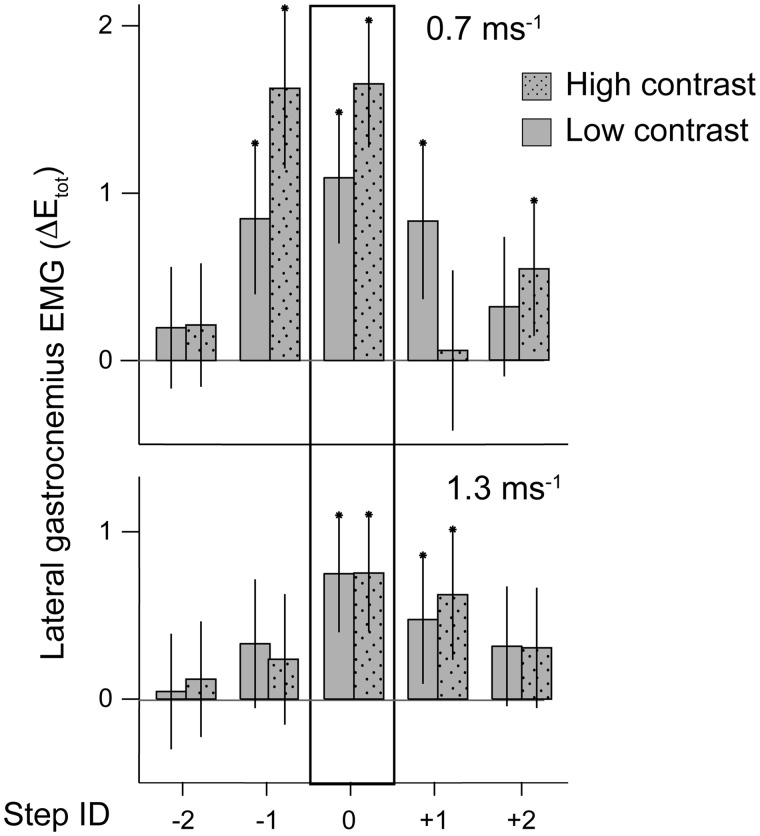
Birds are a diverse and agile lineage of vertebrates that all use bipedal locomotion for at least part of their life. Thus birds provide a valuable opportunity to investigate how biomechanics and sensorimotor control are integrated for agile bipedal locomotion. This review summarizes recent work using terrain perturbations to reveal neuromechanical control strategies used by ground birds to achieve robust, stable, and agile running. Early experiments in running guinea fowl aimed to reveal the immediate intrinsic mechanical response to an unexpected drop ("pothole") in terrain. When navigating the pothole, guinea fowl experience large changes in leg posture in the perturbed step, which correlates strongly with leg loading and perturbation recovery. Analysis of simple theoretical models of running has further confirmed the crucial role of swing-leg trajectory control for regulating foot contact timing and leg loading in uneven terrain. Coupling between body and leg dynamics results in an inherent trade-off in swing leg retraction rate for fall avoidance versus injury avoidance. Fast leg retraction minimizes injury risk, but slow leg retraction minimizes fall risk. Subsequent experiments have investigated how birds optimize their control strategies depending on the type of perturbation (pothole, step, obstacle), visibility of terrain, and with ample practice negotiating terrain features. Birds use several control strategies consistently across terrain contexts: (1) independent control of leg angular cycling and leg length actuation, which facilitates dynamic stability through simple control mechanisms, (2) feedforward regulation of leg cycling rate, which tunes foot-contact timing to maintain consistent leg loading in uneven terrain (minimizing fall and injury risks), (3) load-dependent muscle actuation, which rapidly adjusts stance push-off and stabilizes body mechanical energy, and (4) multi-step recovery strategies that allow body dynamics to transiently vary while tightly regulating leg loading to minimize risks of fall and injury. In future work, it will be interesting to investigate the learning and adaptation processes that allow animals to adjust neuromechanical control mechanisms over short and long timescales.
Figures
References
-
- Abourachid A, Höfling E.. 2012. The legs: a key to bird evolutionary success. J Ornithol 153:193–8.
-
- Alexander RM. 2002. Stability and manoeuvrability of terrestrial vertebrates. Integr Comp Biol 42:158–64. - PubMed
-
- Askew GN, Marsh RL.. 1998. Optimal shortening velocity (V/Vmax) of skeletal muscle during cyclical contractions: length-force effects and velocity-dependent activation and deactivation. J Exp Biol 201:1527–40. - PubMed
-
- Biewener AA, Baudinette RV.. 1995. In vivo muscle force and elastic energy storage during steady-speed hopping of tammar wallabies (Macropus eugenii). J Exp Biol 198:1829–41. - PubMed
-
- Birn-Jeffery AV, Daley MA.. 2012. Birds achieve high robustness in uneven terrain through active control of landing conditions. J Exp Biol 215:2117–27. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
