

Multiple Host Factors Interact with the Hypervariable Domain of Chikungunya Virus nsP3 and Determine Viral Replication in Cell-Specific Mode
- PMID: 29899097
- PMCID: PMC6069204
- DOI: 10.1128/JVI.00838-18
Multiple Host Factors Interact with the Hypervariable Domain of Chikungunya Virus nsP3 and Determine Viral Replication in Cell-Specific Mode
Abstract
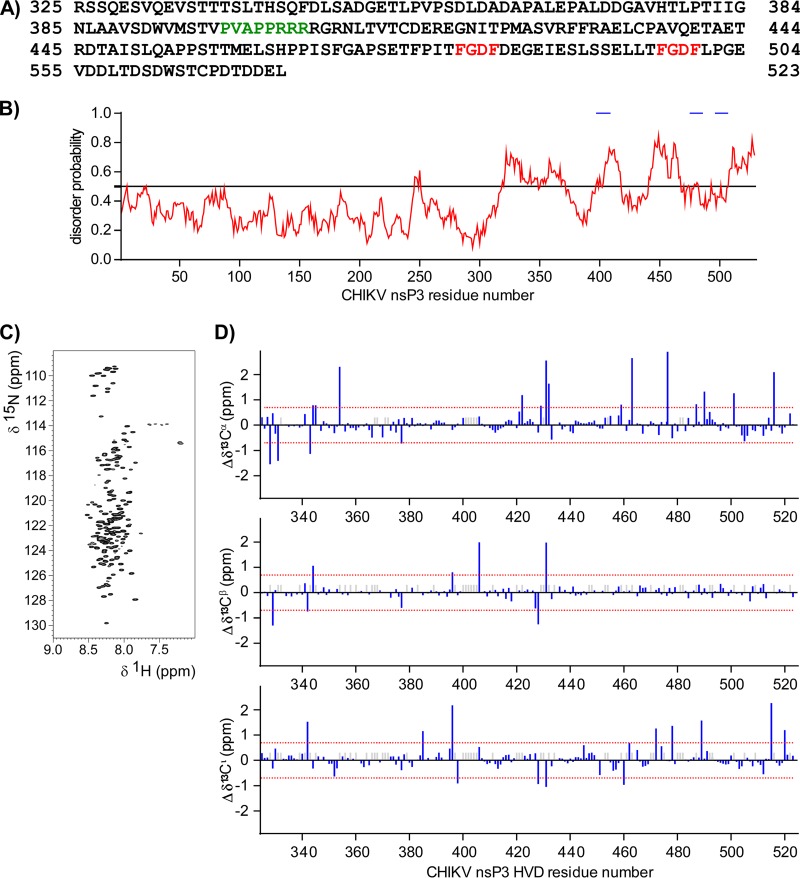
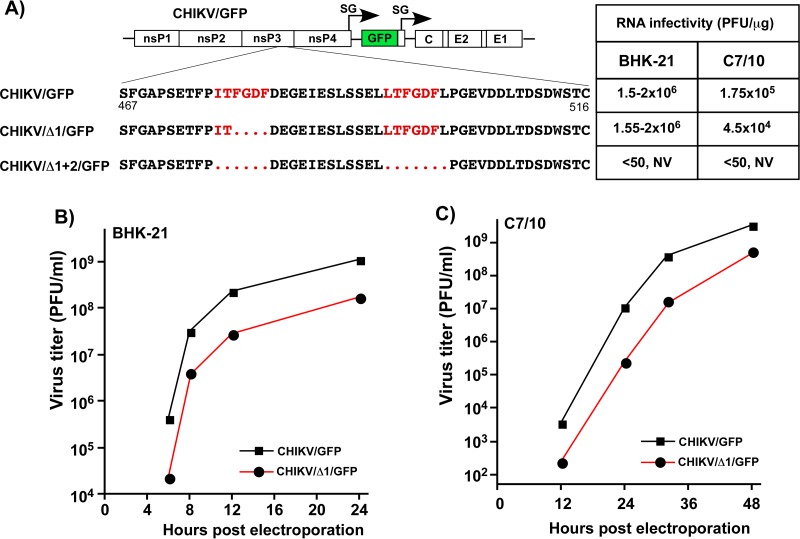
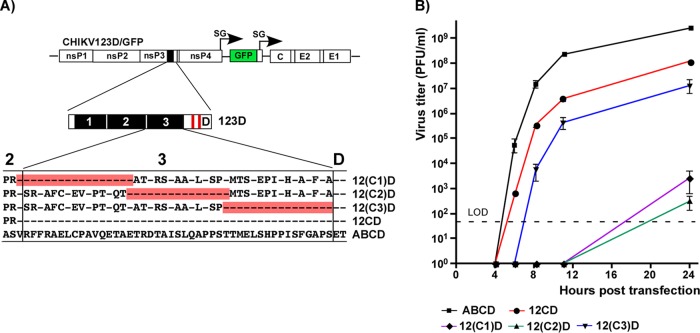
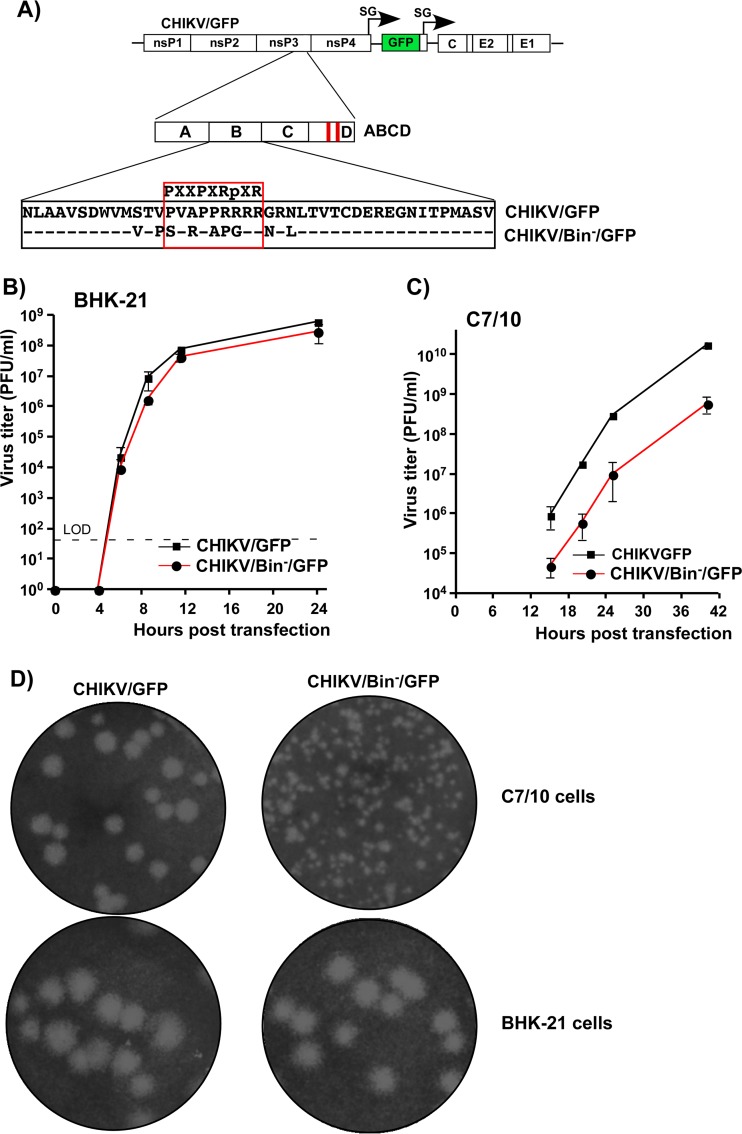
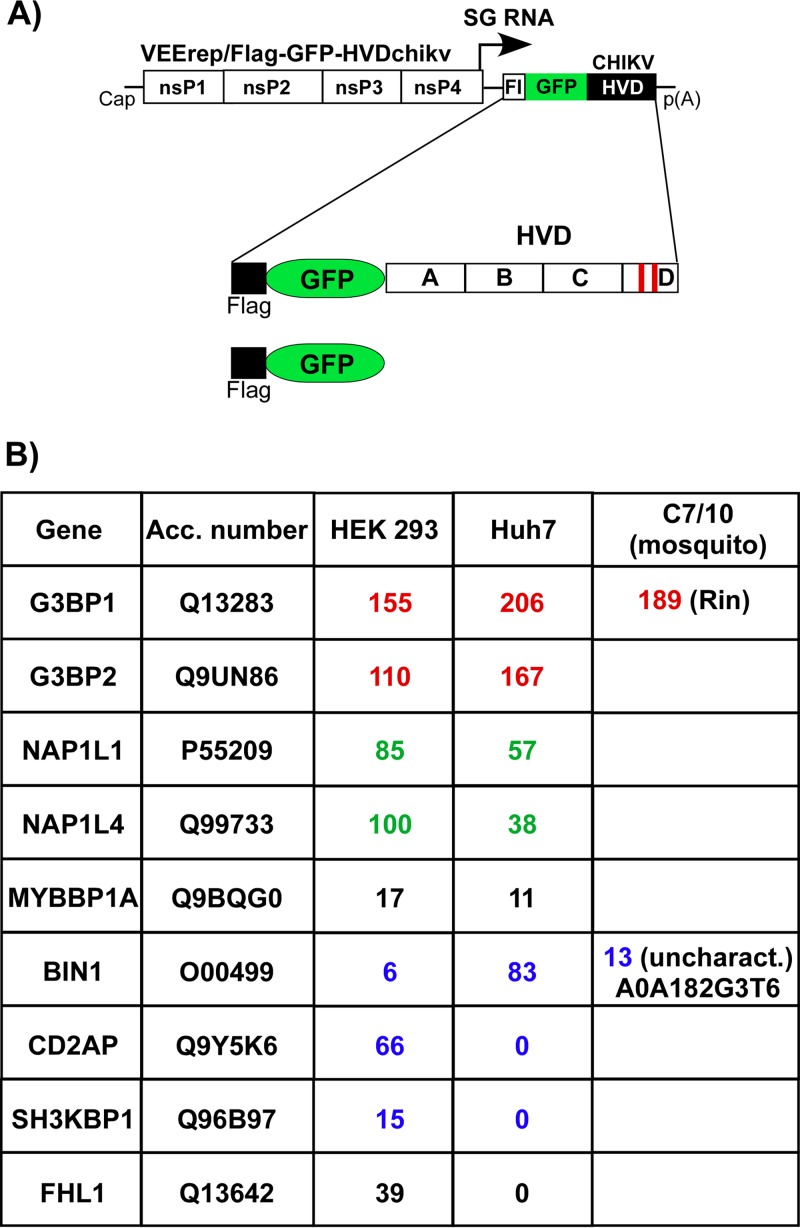
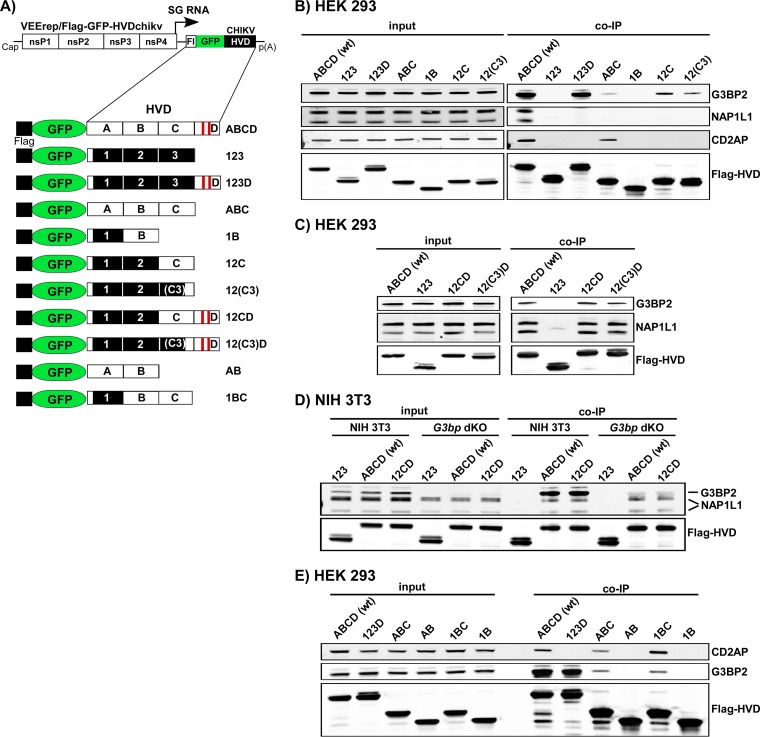
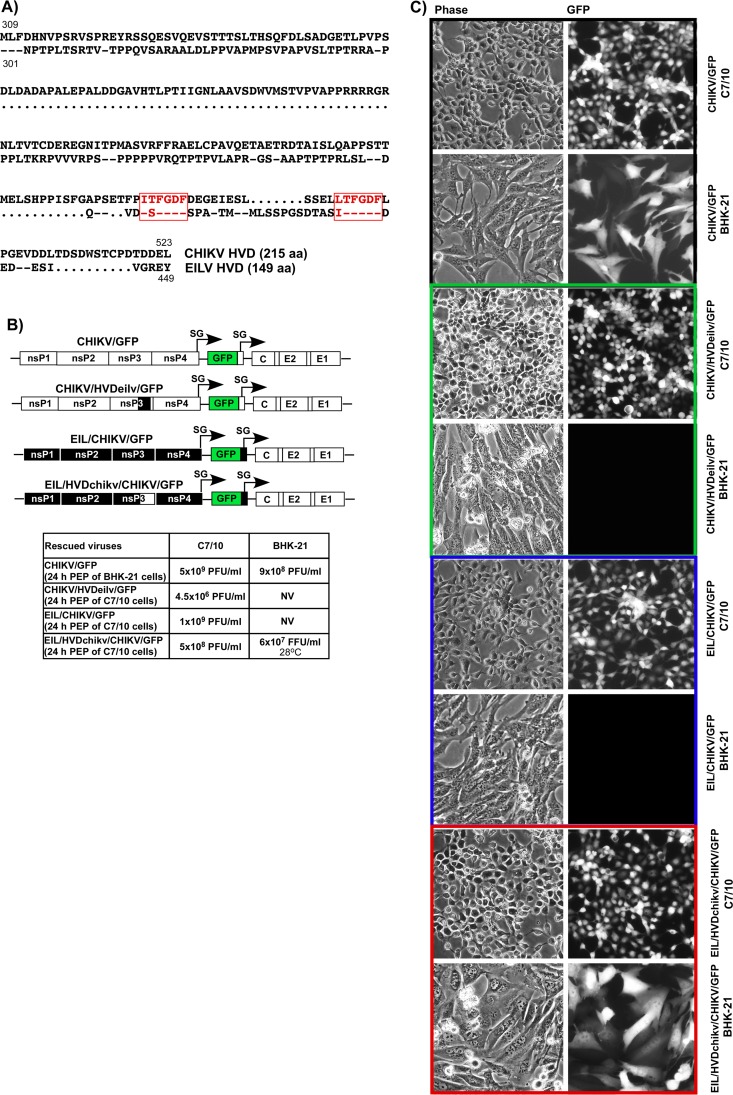
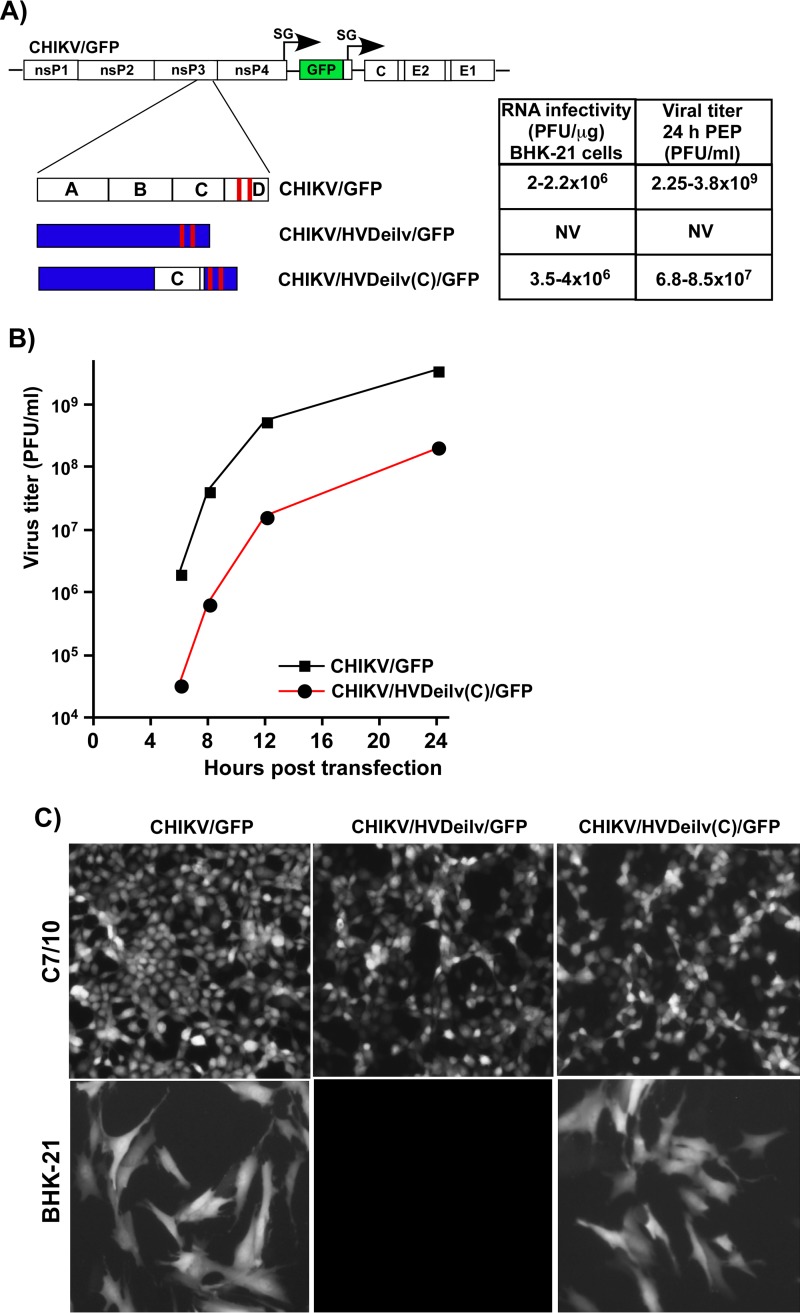
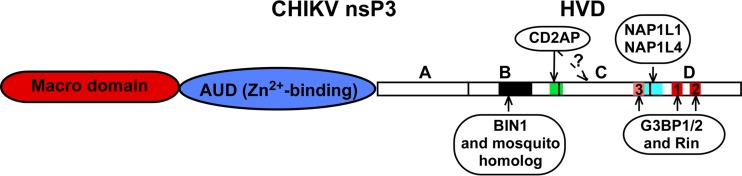
Alphaviruses are widely distributed in both hemispheres and circulate between mosquitoes and amplifying vertebrate hosts. Geographically separated alphaviruses have adapted to replication in particular organisms. The accumulating data suggest that this adaptation is determined not only by changes in their glycoproteins but also by the amino acid sequence of the hypervariable domain (HVD) of the alphavirus nsP3 protein. We performed a detailed investigation of chikungunya virus (CHIKV) nsP3 HVD interactions with host factors and their roles in viral replication in vertebrate and mosquito cells. The results demonstrate that CHIKV HVD is intrinsically disordered and binds several distinctive cellular proteins. These host factors include two members of the G3BP family and their mosquito homolog Rin, two members of the NAP1 family, and several SH3 domain-containing proteins. Interaction with G3BP proteins or Rin is an absolute requirement for CHIKV replication, although it is insufficient to solely drive it in either vertebrate or mosquito cells. To achieve a detectable level of virus replication, HVD needs to bind members of at least one more protein family in addition to G3BPs. Interaction with NAP1L1 and NAP1L4 plays a more proviral role in vertebrate cells, while binding of SH3 domain-containing proteins to a proline-rich fragment of HVD is more critical for virus replication in the cells of mosquito origin. Modifications of binding sites in CHIKV HVD allow manipulation of the cell specificity of CHIKV replication. Similar changes may be introduced into HVDs of other alphaviruses to alter their replication in particular cells or tissues.IMPORTANCE Alphaviruses utilize a broad spectrum of cellular factors for efficient formation and function of replication complexes (RCs). Our data demonstrate for the first time that the hypervariable domain (HVD) of chikungunya virus nonstructural protein 3 (nsP3) is intrinsically disordered. It binds at least 3 families of cellular proteins, which play an indispensable role in viral RNA replication. The proteins of each family demonstrate functional redundancy. We provide a detailed map of the binding sites on CHIKV nsP3 HVD and show that mutations in these sites or the replacement of CHIKV HVD by heterologous HVD change cell specificity of viral replication. Such manipulations with alphavirus HVDs open an opportunity for development of new irreversibly attenuated vaccine candidates. To date, the disordered protein fragments have been identified in the nonstructural proteins of many other viruses. They may also interact with a variety of cellular factors that determine critical aspects of virus-host interactions.
Keywords: BIN1; CD2AP; Eilat virus; G3BP1; G3BP2; NAP1L1; alphavirus; chikungunya virus; viral RNA replication; virus-host interactions.
Copyright © 2018 American Society for Microbiology.
Figures
References
-
- Griffin DE. 2001. Alphaviruses, p 917–962. In Knipe DM, Howley PM, Griffin DE, Lamb RA, Martin MA, Roizman B, Straus SE (ed), Fields virology, 4th ed Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Weaver SC, Frolov I. 2005. Togaviruses, p 1010–1024. In Mahy BWJ, ter Meulen V (ed), Virology, vol 2 Hodder Arnold, Salisbury, United Kingdom.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
