

Substrate deformations induce directed keratinocyte migration
- PMID: 29899159
- PMCID: PMC6030620
- DOI: 10.1098/rsif.2018.0133
Substrate deformations induce directed keratinocyte migration
Abstract
Cell migration is an essential part of many (patho)physiological processes, including keratinocyte re-epithelialization of healing wounds. Physical forces and mechanical cues from the wound bed (in addition to biochemical signals) may also play an important role in the healing process. Previously, we explored this possibility and found that polyacrylamide (PA) gel stiffness affected human keratinocyte behaviour and that mechanical deformations in soft (approx. 1.2 kPa) PA gels produced by neighbouring cells appeared to influence the process of de novo epithelial sheet formation. To clearly demonstrate that keratinocytes do respond to such deformations, we conducted a series of experiments where we observed the response of single keratinocytes to a prescribed local substrate deformation that mimicked a neighbouring cell or evolving multicellular aggregate via a servo-controlled microneedle. We also examined the effect of adding either Y27632 or blebbistatin on cell response. Our results indicate that keratinocytes do sense and respond to mechanical signals comparable to those that originate from substrate deformations imposed by neighbouring cells, a finding that could have important implications for the process of keratinocyte re-epithelialization that takes place during wound healing. Furthermore, the Rho/ROCK pathway and the engagement of NM II are both essential to substrate deformation-directed keratinocyte migration.
Keywords: actomyosin; mechanobiology; mechanosensing; non-muscle myosin II; polyacrylamide gel.
© 2018 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures
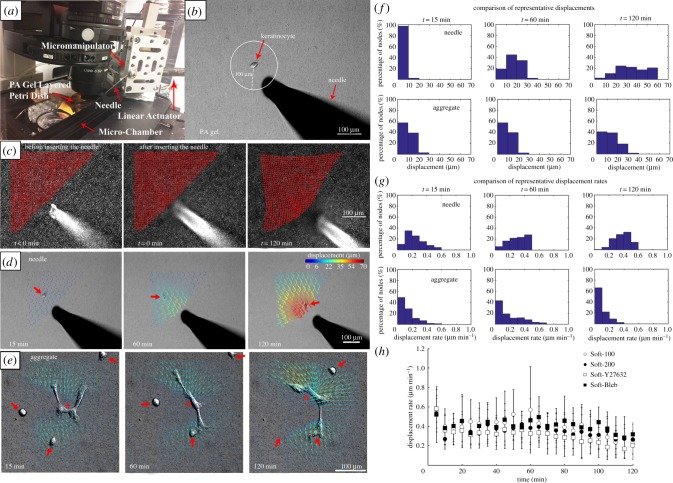

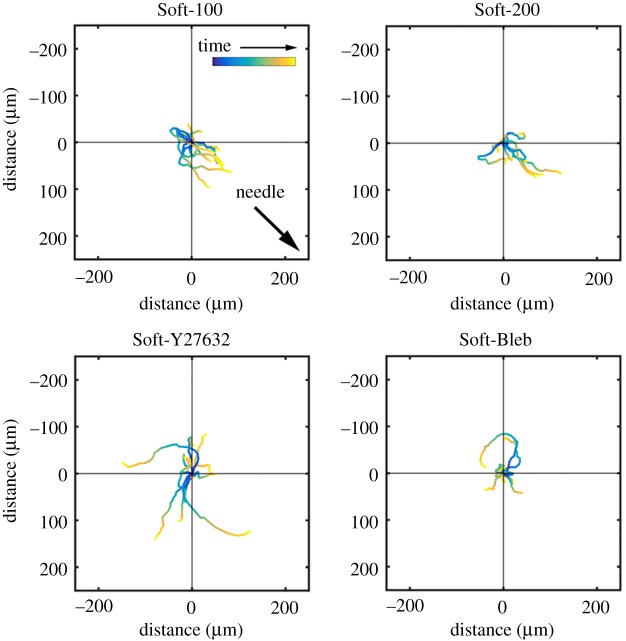
 , the position vector of the cell due to cell migration effects alone (see equation (2.2)). Each cell path is colour-coded to show the relative time along the path from its beginning at the origin (dark blue) to the final tracked position (yellow). The majority of cells moved toward the displacing needle for needles nominally positioned either 100 or 200 µm from the cell centroid at the start of the experiment (i.e. Soft-100 and Soft-200, respectively). Exposure to the Rho kinase inhibitor Y27632 or the actin–myosin inhibitor blebbistatin interrupted this directed migration. (Online version in colour.)
, the position vector of the cell due to cell migration effects alone (see equation (2.2)). Each cell path is colour-coded to show the relative time along the path from its beginning at the origin (dark blue) to the final tracked position (yellow). The majority of cells moved toward the displacing needle for needles nominally positioned either 100 or 200 µm from the cell centroid at the start of the experiment (i.e. Soft-100 and Soft-200, respectively). Exposure to the Rho kinase inhibitor Y27632 or the actin–myosin inhibitor blebbistatin interrupted this directed migration. (Online version in colour.)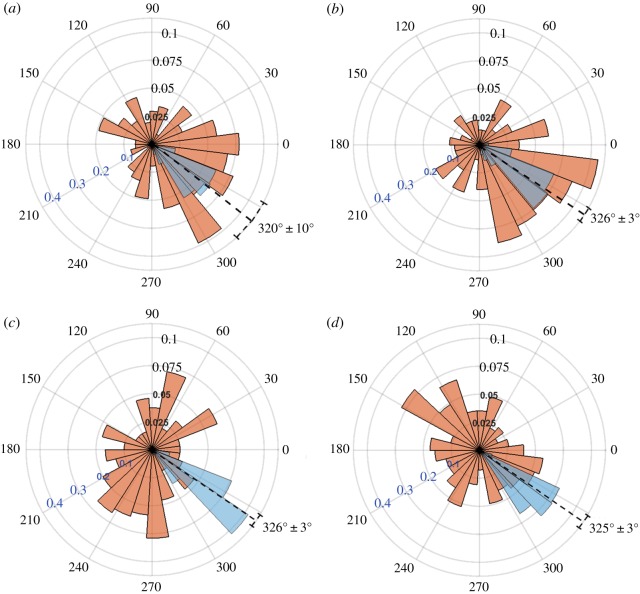
 (red) and substrate displacement
(red) and substrate displacement  (blue) for all 10 replicates of the (a) Soft-100, (b) Soft-200, (c) Soft-Y27632 and (d) Soft-Bleb experiments. Each distribution is normalized by the total number of observations with the black radial text associated with
(blue) for all 10 replicates of the (a) Soft-100, (b) Soft-200, (c) Soft-Y27632 and (d) Soft-Bleb experiments. Each distribution is normalized by the total number of observations with the black radial text associated with  and the blue radial text associated with
and the blue radial text associated with  . The dashed line shows the mean and standard deviation of the angle of needle movement. For all four conditions, the displacements of the substrate aligned with the direction of needle movement. In addition, for Soft-100 and Soft-200, V-tests indicate that the direction of cell movement is coincident with that of the needle-induced substrate deformations, but not for soft-Y27632 or Soft-Bleb (p < 0.001). (Online version in colour.)
. The dashed line shows the mean and standard deviation of the angle of needle movement. For all four conditions, the displacements of the substrate aligned with the direction of needle movement. In addition, for Soft-100 and Soft-200, V-tests indicate that the direction of cell movement is coincident with that of the needle-induced substrate deformations, but not for soft-Y27632 or Soft-Bleb (p < 0.001). (Online version in colour.)References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources