

Avian tail ontogeny, pygostyle formation, and interpretation of juvenile Mesozoic specimens
- PMID: 29899503
- PMCID: PMC5997987
- DOI: 10.1038/s41598-018-27336-x
Avian tail ontogeny, pygostyle formation, and interpretation of juvenile Mesozoic specimens
Abstract
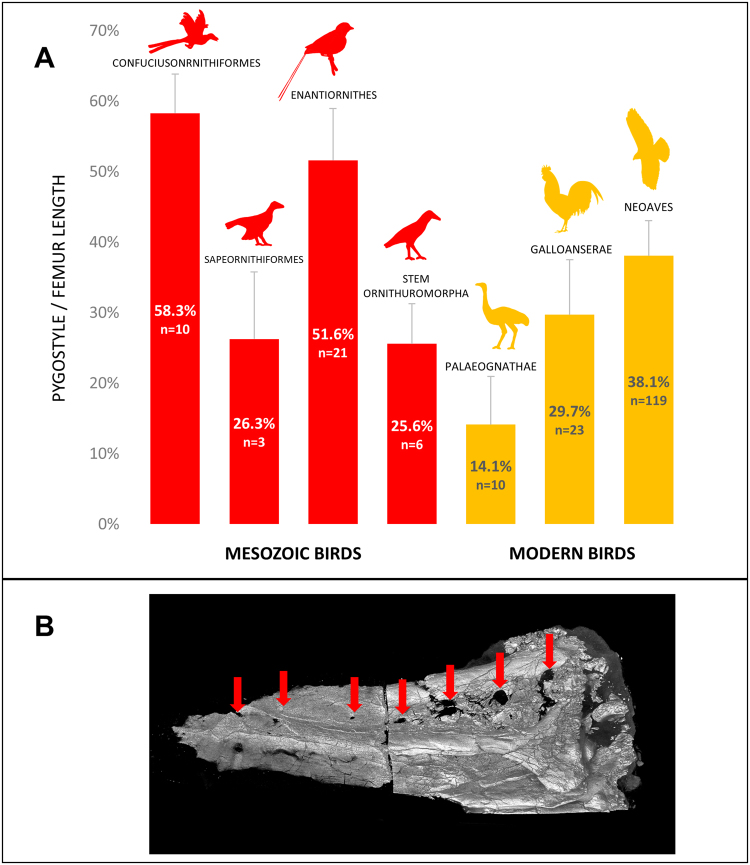
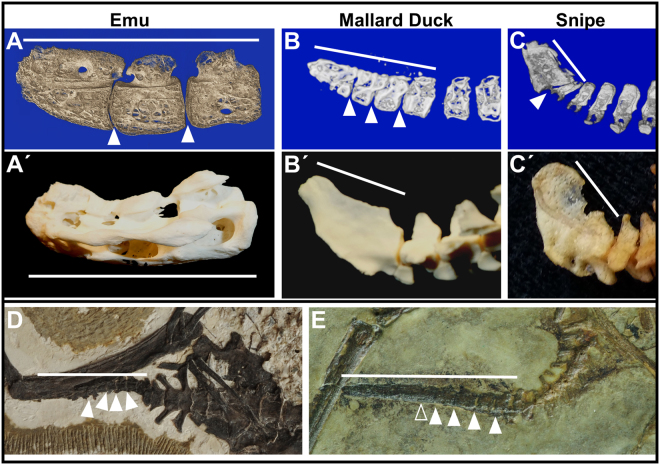
The avian tail played a critical role in the evolutionary transition from long- to short-tailed birds, yet its ontogeny in extant birds has largely been ignored. This deficit has hampered efforts to effectively identify intermediate species during the Mesozoic transition to short tails. Here we show that fusion of distal vertebrae into the pygostyle structure does not occur in extant birds until near skeletal maturity, and mineralization of vertebral processes also occurs long after hatching. Evidence for post-hatching pygostyle formation is also demonstrated in two Cretaceous specimens, a juvenile enantiornithine and a subadult basal ornithuromorph. These findings call for reinterpretations of Zhongornis haoae, a Cretaceous bird hypothesized to be an intermediate in the long- to short-tailed bird transition, and of the recently discovered coelurosaur tail embedded in amber. Zhongornis, as a juvenile, may not yet have formed a pygostyle, and the amber-embedded tail specimen is reinterpreted as possibly avian. Analyses of relative pygostyle lengths in extant and Cretaceous birds suggests the number of vertebrae incorporated into the pygostyle has varied considerably, further complicating the interpretation of potential transitional species. In addition, this analysis of avian tail development reveals the generation and loss of intervertebral discs in the pygostyle, vertebral bodies derived from different kinds of cartilage, and alternative modes of caudal vertebral process morphogenesis in birds. These findings demonstrate that avian tail ontogeny is a crucial parameter specifically for the interpretation of Mesozoic specimens, and generally for insights into vertebrae formation.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
An Enantiornithine with a Fan-Shaped Tail, and the Evolution of the Rectricial Complex in Early Birds.Curr Biol. 2016 Jan 11;26(1):114-9. doi: 10.1016/j.cub.2015.11.036. Epub 2015 Dec 31. Curr Biol. 2016. PMID: 26748849
-
A bizarre Early Cretaceous enantiornithine bird with unique crural feathers and an ornithuromorph plough-shaped pygostyle.Nat Commun. 2017 Jan 31;8:14141. doi: 10.1038/ncomms14141. Nat Commun. 2017. PMID: 28139644 Free PMC article.
-
Coevolution of caudal skeleton and tail feathers in birds.J Morphol. 2014 Dec;275(12):1431-40. doi: 10.1002/jmor.20321. Epub 2014 Aug 20. J Morphol. 2014. PMID: 25139752
-
From dinosaurs to birds: a tail of evolution.Evodevo. 2014 Jul 29;5:25. doi: 10.1186/2041-9139-5-25. eCollection 2014. Evodevo. 2014. PMID: 25621146 Free PMC article. Review.
-
From extant to extinct: locomotor ontogeny and the evolution of avian flight.Trends Ecol Evol. 2012 May;27(5):296-305. doi: 10.1016/j.tree.2011.12.003. Epub 2012 Feb 1. Trends Ecol Evol. 2012. PMID: 22304966 Review.
Cited by
-
Nonpathological inflammation drives the development of an avian flight adaptation.Proc Natl Acad Sci U S A. 2023 May 9;120(19):e2219757120. doi: 10.1073/pnas.2219757120. Epub 2023 May 1. Proc Natl Acad Sci U S A. 2023. PMID: 37126698 Free PMC article.
-
Distal spinal nerve development and divergence of avian groups.Sci Rep. 2020 Apr 14;10(1):6303. doi: 10.1038/s41598-020-63264-5. Sci Rep. 2020. PMID: 32286419 Free PMC article.
-
Palaeontological evidence reveals convergent evolution of intervertebral joint types in amniotes.Sci Rep. 2020 Aug 24;10(1):14106. doi: 10.1038/s41598-020-70751-2. Sci Rep. 2020. PMID: 32839497 Free PMC article.
-
A new confuciusornithid bird with a secondary epiphyseal ossification reveals phylogenetic changes in confuciusornithid flight mode.Commun Biol. 2022 Dec 21;5(1):1398. doi: 10.1038/s42003-022-04316-6. Commun Biol. 2022. PMID: 36543908 Free PMC article.
-
Ontogeny of the anuran urostyle and the developmental context of evolutionary novelty.Proc Natl Acad Sci U S A. 2020 Feb 11;117(6):3034-3044. doi: 10.1073/pnas.1917506117. Epub 2020 Jan 27. Proc Natl Acad Sci U S A. 2020. PMID: 31988131 Free PMC article.
References
-
- Chatterjee, S. The Rise of Birds: 225 Million Years of Evolution, (Johns Hopkins University Press) (1997).
-
- Chiappe, L. M. & Witmer, L. M. Mesozoic Birds: Above the Heads of Dinosaurs, (Berkeley, Los Angeles, and London: University of California Press) (2002).
-
- Huxley TH. On the animals which are most nearly intermediate between birds and reptiles. Geol. Mag. 1868;5:357–365. doi: 10.1017/S0016756800207784. - DOI
-
- Huxley TH. Further Evidence of the Affinity between the Dinosaurian Reptiles and Birds. Quarterly Journal of the Geological Society. 1870;26:12–31. doi: 10.1144/GSL.JGS.1870.026.01-02.08. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
