

Human primitive brain displays negative mitochondrial-nuclear expression correlation of respiratory genes
- PMID: 29903725
- PMCID: PMC6028125
- DOI: 10.1101/gr.226324.117
Human primitive brain displays negative mitochondrial-nuclear expression correlation of respiratory genes
Erratum in
-
Corrigendum: Human primitive brain displays negative mitochondrial-nuclear expression correlation of respiratory genes.Genome Res. 2022 Aug 25;32(8):1626. doi: 10.1101/gr.277125.122. Genome Res. 2022. PMID: 39074347 Free PMC article. No abstract available.
Abstract
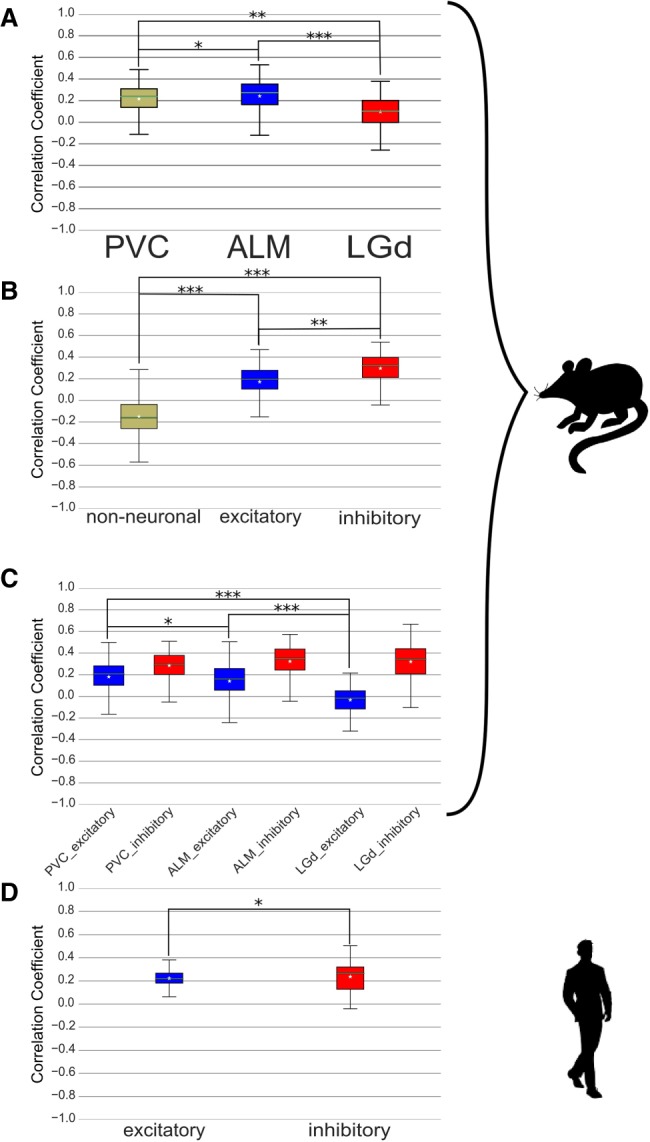
Oxidative phosphorylation (OXPHOS), a fundamental energy source in all human tissues, requires interactions between mitochondrial (mtDNA)- and nuclear (nDNA)-encoded protein subunits. Although such interactions are fundamental to OXPHOS, bi-genomic coregulation is poorly understood. To address this question, we analyzed ∼8500 RNA-seq experiments from 48 human body sites. Despite well-known variation in mitochondrial activity, quantity, and morphology, we found overall positive mtDNA-nDNA OXPHOS genes' co-expression across human tissues. Nevertheless, negative mtDNA-nDNA gene expression correlation was identified in the hypothalamus, basal ganglia, and amygdala (subcortical brain regions, collectively termed the "primitive" brain). Single-cell RNA-seq analysis of mouse and human brains revealed that this phenomenon is evolutionarily conserved, and both are influenced by brain cell types (involving excitatory/inhibitory neurons and nonneuronal cells) and by their spatial brain location. As the "primitive" brain is highly oxidative, we hypothesized that such negative mtDNA-nDNA co-expression likely controls for the high mtDNA transcript levels, which enforce tight OXPHOS regulation, rather than rewiring toward glycolysis. Accordingly, we found "primitive" brain-specific up-regulation of lactate dehydrogenase B (LDHB), which associates with high OXPHOS activity, at the expense of LDHA, which promotes glycolysis. Analyses of co-expression, DNase-seq, and ChIP-seq experiments revealed candidate RNA-binding proteins and CEBPB as the best regulatory candidates to explain these phenomena. Finally, cross-tissue expression analysis unearthed tissue-dependent splice variants and OXPHOS subunit paralogs and allowed revising the list of canonical OXPHOS transcripts. Taken together, our analysis provides a comprehensive view of mito-nuclear gene co-expression across human tissues and provides overall insights into the bi-genomic regulation of mitochondrial activities.
© 2018 Barshad et al.; Published by Cold Spring Harbor Laboratory Press.
Figures






References
-
- Azevedo FA, Carvalho LR, Grinberg LT, Farfel JM, Ferretti RE, Leite RE, Lent R, Herculano-Houzel S. 2009. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J Comp Neurol 513: 532–541. - PubMed
-
- Barton RA, Venditti C. 2014. Rapid evolution of the cerebellum in humans and other great apes. Curr Biol 24: 2440–2444. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous