

VRTN is Required for the Development of Thoracic Vertebrae in Mammals
- PMID: 29904281
- PMCID: PMC6001657
- DOI: 10.7150/ijbs.23815
VRTN is Required for the Development of Thoracic Vertebrae in Mammals
Abstract
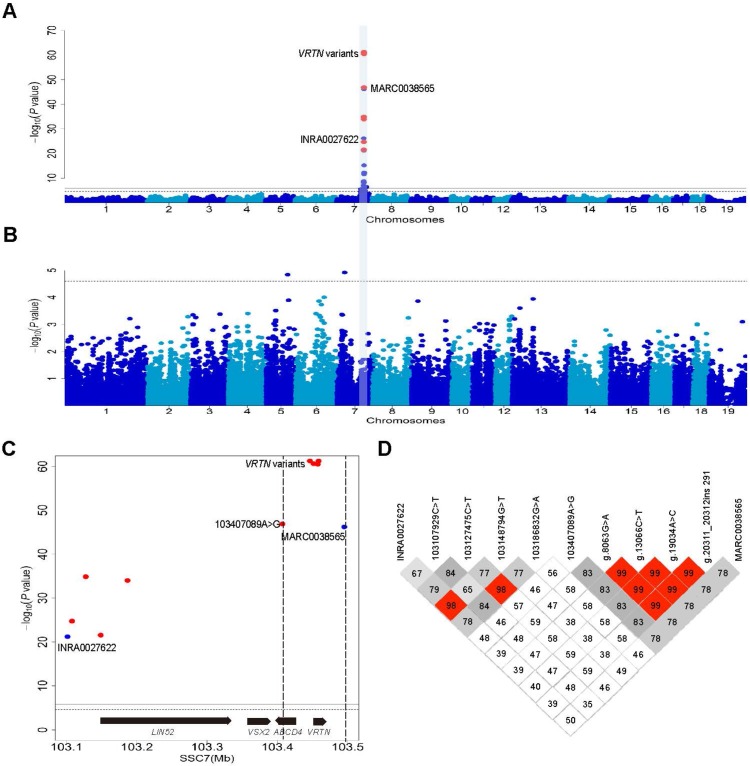
Vertnin (VRTN) variants are associated with thoracic vertebral number (TVN) in pigs. However, the biological function of VRTN remains poorly understood. Here we first conducted a range of experiments to demonstrate that VRTN is a responsible gene for TVN and two causative variants in the regulatory region of VRTN additively regulate TVN. Then, we show that VRTN is a novel DNA-binding transcription factor as it localizes exclusively in the nucleus, binds to DNA on a genome-wide scale and regulates the transcription of a set of genes that harbor VRTN binding motifs. Next, we illustrate that VRTN is essential for the development of thoracic vertebrae. Vrtn-null embryos display somitogenesis defect with the failure of axial rotation and fewer somites at the thoracic somite stage. Half of Vrtn heterozygous mice show abnormal spinal development with fewer thoracic vertebrae and ribs than their wild-type littermates. Lastly, we reveal that VRTN could modulate somite segmentation via the Notch signaling pathway. The findings advance our understanding of the mechanisms underlying the development of thoracic vertebrate in mammals, and VRTN causative variants provide a robust tool to improve pork production by selecting the alleles increasing the number of thoracic vertebrae and ribs.
Keywords: VRTN; somitogenesis; thoracic vertebrae; transcription factor.
Conflict of interest statement
Competing Interests: The authors have declared that no competing interest exists.
Figures




Similar articles
-
Possible introgression of the VRTN mutation increasing vertebral number, carcass length and teat number from Chinese pigs into European pigs.Sci Rep. 2016 Jan 19;6:19240. doi: 10.1038/srep19240. Sci Rep. 2016. PMID: 26781738 Free PMC article.
-
A further look at porcine chromosome 7 reveals VRTN variants associated with vertebral number in Chinese and Western pigs.PLoS One. 2013 Apr 24;8(4):e62534. doi: 10.1371/journal.pone.0062534. Print 2013. PLoS One. 2013. PMID: 23638110 Free PMC article.
-
Effect of VRTN gene polymorphisms on Duroc pig production and carcass traits, and their genetic relationships.Anim Sci J. 2015 Feb;86(2):125-31. doi: 10.1111/asj.12260. Epub 2014 Sep 3. Anim Sci J. 2015. PMID: 25187328
-
The many roles of Notch signaling during vertebrate somitogenesis.Semin Cell Dev Biol. 2016 Jan;49:68-75. doi: 10.1016/j.semcdb.2014.11.010. Epub 2014 Dec 4. Semin Cell Dev Biol. 2016. PMID: 25483003 Review.
-
The vertebrate segmentation clock and its role in skeletal birth defects.Birth Defects Res C Embryo Today. 2007 Jun;81(2):121-33. doi: 10.1002/bdrc.20090. Birth Defects Res C Embryo Today. 2007. PMID: 17600784 Review.
Cited by
-
Transcriptomics and Selection Pressure Analysis Reveals the Influence Mechanism of PLIN1 Protein on the Development of Small Size in Min Pigs.Int J Mol Sci. 2023 Feb 15;24(4):3947. doi: 10.3390/ijms24043947. Int J Mol Sci. 2023. PMID: 36835359 Free PMC article.
-
Genetic dissection and genomic prediction for pork cuts and carcass morphology traits in pig.J Anim Sci Biotechnol. 2023 Sep 3;14(1):116. doi: 10.1186/s40104-023-00914-4. J Anim Sci Biotechnol. 2023. PMID: 37660101 Free PMC article.
-
Genome-Wide Association Analysis Identified BMPR1A as a Novel Candidate Gene Affecting the Number of Thoracic Vertebrae in a Large White × Minzhu Intercross Pig Population.Animals (Basel). 2020 Nov 22;10(11):2186. doi: 10.3390/ani10112186. Animals (Basel). 2020. PMID: 33266466 Free PMC article.
-
A Whole Genome Sequencing-Based Genome-Wide Association Study Reveals the Potential Associations of Teat Number in Qingping Pigs.Animals (Basel). 2022 Apr 20;12(9):1057. doi: 10.3390/ani12091057. Animals (Basel). 2022. PMID: 35565484 Free PMC article.
-
Detection and Analysis of Genes Affecting the Number of Thoracic Vertebrae in Licha Black Pigs.Genes (Basel). 2024 Apr 10;15(4):477. doi: 10.3390/genes15040477. Genes (Basel). 2024. PMID: 38674411 Free PMC article.
References
-
- Sparrow DB, Chapman G, Turnpenny PD, Dunwoodie SL. Disruption of the somitic molecular clock causes abnormal vertebral segmentation. Birth Defects Research. 2007;81:93–110. - PubMed
-
- Narita Y, Kuratani S. Evolution of the vertebral formulae in mammals: a perspective on developmental constraints. Journal of Experimental Zoology Part B, Molecular and Developmental Evolution. 2005;304:91–106. - PubMed
-
- Asher RJ, Lin KH, Kardjilov N, Hautier L. Variability and constraint in the mammalian vertebral column. Journal of Evolutionary Biology. 2011;24:1080–90. - PubMed
-
- Zhang ZG LB, Chen XH. Pig breeds in China. Shanghai Scientific and Technical Publisher, Shang, China; 1986.
-
- Borchers N. RN, Kalm E. The number of ribs and vertebrae in a Pie´train cross: variation, heritability and effects on performance traits. Jouranl of Animal Breeding and Genetics. 2004;121:392–403.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
